Ergosterol is as ubiquitous in mushrooms as cholesterol is in humans. It is formed by an almost identical metabolic process, the mevalonate pathway. When mushrooms are exposed to ultraviolet light, ergosterol is converted to ergocalciferol, or vitamin D2. Once ingested, vitamin D2 is converted to calcidiol in the liver and eventually calcitriol, our active vitamin D3, in the kidneys. There is clinical evidence demonstrating that ergocalciferol-D2, although not as bioavailable as calcitriol, could improve vitamin D deficiency in humans (1,2,3).
Mushrooms are an excellent source of vitamin D2, particularly if they have been exposed to ample UV light (4). Researchers found that when people consumed chanterelle mushrooms containing 14 mcg of ergocalciferol with their lunch, that their serum vitamin D levels improved as much as the group who was receiving ergocalciferol supplementation (5). However, another study found that vitamin D levels decreased significantly after cooking mushrooms and serum concentrations of vitamin D after consumption of 100 g fresh sliced cooked mushrooms for 16 weeks did not show significant improvement in serum vitamin D levels. This was, notably, in a prediabetic, overweight population (6). In healthy young adults, serum vitamin D levels did improve after consumption of vitamin D2-enhanced button mushrooms via UV-B irradiation and to a similar degree as with a vitamin D2 supplement (7).
The benefits of optimal serum vitamin D levels include and are not limited to:
Bone health, kidney health, immune modulation, calcium homeostasis, and cardiovascular health (8).
Vaes AMM, Tieland M, de Regt MF, Wittwer J, van Loon LJC, de Groot LCPGM. Dose-response effects of supplementation with calcifediol on serum 25-hydroxyvitamin D status and its metabolites: A randomized controlled trial in older adults. Clin Nutr. 2018;37(3):808-814. doi:10.1016/j.clnu.2017.03.029
Martineau AR, Thummel KE, Wang Z, et al. Differential Effects of Oral Boluses of Vitamin D2 vs Vitamin D3 on Vitamin D Metabolism: A Randomized Controlled Trial. J Clin Endocrinol Metab. 2019;104(12):5831-5839. doi:10.1210/jc.2019-00207
Seijo M, Mastaglia S, Brito G, Somoza J, Oliveri B. Es equivalente la suplementación diaria con vitamina D2 o vitamina D3 en adultos mayores? [Is daily supplementation with vitamin D2 equivalent to daily supplementation with vitamin D3 in the elderly?]. Medicina (B Aires). 2012;72(3):195-200.
Kamweru PK, Tindibale EL. Vitamin D and Vitamin D from Ultraviolet-Irradiated Mushrooms (Review). Int J Med Mushrooms. 2016;18(3):205-214. doi:10.1615/IntJMedMushrooms.v18.i3.30
Outila TA, Mattila PH, Piironen VI, Lamberg-Allardt CJ. Bioavailability of vitamin D from wild edible mushrooms (Cantharellus tubaeformis) as measured with a human bioassay. Am J Clin Nutr. 1999;69(1):95-98. doi:10.1093/ajcn/69.1.95
Mehrotra A, Calvo MS, Beelman RB, et al. Bioavailability of vitamin D2 from enriched mushrooms in prediabetic adults: a randomized controlled trial. Eur J Clin Nutr. 2014;68(10):1154-1160. doi:10.1038/ejcn.2014.157
Urbain P, Singler F, Ihorst G, Biesalski HK, Bertz H. Bioavailability of vitamin D₂ from UV-B-irradiated button mushrooms in healthy adults deficient in serum 25-hydroxyvitamin D: a randomized controlled trial. Eur J Clin Nutr. 2011;65(8):965-971. doi:10.1038/ejcn.2011.53
Charoenngam N, Holick MF. Immunologic Effects of Vitamin D on Human Health and Disease. Nutrients. 2020;12(7):2097. Published 2020 Jul 15. doi:10.3390/nu12072097
Terpenes are fat- and alcohol-soluble compounds that come in many variations – monoterpenes, sesquiterpenes, diterpenes and triterpenes. The prefix indicates the number of carbons in the molecule: 10, 15, 20, and 30, respectively. As the terpene becomes bigger, it also becomes heavier and less volatile. The mushroom smell many are familiar with is in part due to low-molecular weight molecules that make their way into the nose – the monoterpenes and sesquiterpenes. These lower-weight chemicals act as attractants and deterrents to insects and other invertebrates, acting as both pheromones and defense chemicals. The most well-known terpenes in fungi are both found in lion’s mane. Erinicines, a group of diterpenoids, have nootropic properties and are found in lion’s mane mycelium, while the terpene hericenones, which are also nootropic, are found in lion’s mane fruiting body.
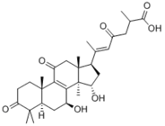
As the mushroom matures from primordia to fruiting body, an enzyme stimulates the synthesis of triterpenes. Because of this, there are significantly more triterpenes in the mushroom compared to the mycelium and primordia. Triterpenes in mushrooms are lanostane triterpenes, and can be referred to as “lanostanoids”. The most well-known triterpenes found in mushrooms are ganoderic acids, found in various species of reishi. These terpenes are major compounds of interest in cancer research because they have cytotoxic activity in many cancer cell lines; they also demonstrate specific antiviral activity, and are helpful in the treatment of hypersensitivity reactions.
Ganoderic acid, a triterpene isolated from Ganoderma spp.
Anti-neoplastic
There are a significant number of in vitro and in vivo animal studies demonstrating direct anti-neoplastic activity of triterpenes. These molecules are cytotoxic to many cancer cell lines, as well as instigators of an antineoplastic immune response (1,2,3,4,5,6).
Anti-viral
Mushroom-derived triterpenes and sesquiterpenes have specific antiviral activity through the inhibition of the enzyme neuraminidase, which viruses use to leave one cell to enter another (7,8). In addition to anti-neuraminidase activity, triterpenes stimulate an immune response that preferences cytotoxic CD8 T cells, a major component of adaptive antiviral immunity (9).
Anti-allergic
Triterpenes may be very beneficial in the treatment of hypersensitivity reactions including allergy, dermatitis and rhinitis. Ganoderic acids from reishi inhibit histamine release from mast cells, which can moderate the severity of allergic reactions (10). Ganoderic acids have also been shown to induce the release of the cytokines IL-10 and IFN-𝛾, suppressing TH2 responses in favor of TH1, and moving the immune system away from allergic responses (11).
Neuroprotective
Erinacines and hericenones, diterpenes found in lion’s mane mycelium and fruiting body, respectively, both have neuroprotective activity (12). Both molecules have demonstrated neurogenesis via nerve growth factor synthase activation, and erinacines improve plaque formation in mouse models of Alzheimer’s disease while increasing hippocampal neurogenesis (12).
Absorption
Only 10% of terpenes found in fungi are bioavailable with oral consumption, potentially due to first-pass metabolism in the liver and metabolism by intestinal bacteria (13). Raw mushrooms offer particularly poor bioavailability, as terpenes remain trapped inside fungal cells, while mushroom extracts offer improved absorption. Taking terpene-rich extracts with food impairs absorption; for best results, take terpene-rich mushrooms as extracts on an empty stomach (14). To learn more, check out my blog post on factors impacting the absorption of mushroom constituents.
DIY Extraction of Terpenes
Terpenes are alcohol- and fat-soluble and range from low- to high-molecular weight
Infuse chopped mushrooms into oil and use as a base for lotion, salad dressings or flavorful drizzle
Soak dried and chopped mushrooms in alcohol as part of a double extraction process.
1. Ríos JL. Effects of triterpenes on the immune system. J Ethnopharmacol. 2010;128(1):1-14. doi:10.1016/j.jep.2009.12.045.
2. Grienke U, Kaserer T, Pfluger F, et al. Accessing biological actions of Ganoderma secondary metabolites by in silico profiling. Phytochemistry. 2015;114:114-124. doi:10.1016/j.phytochem.2014.10.010.
3. Bhattarai G, Lee Y-H, Lee N-H, et al. Fomitoside-K from Fomitopsis nigra Induces Apoptosis of Human Oral Squamous Cell Carcinomas (YD-10B) via Mitochondrial Signaling Pathway. Biol Pharm Bull. 2012;35(10):1711-1719. doi:10.1248/bpb.12-00297.
4. Ren G, Liu XY, Zhu HK, Yang SZ, Fu CX. Evaluation of cytotoxic activities of some medicinal polypore fungi from China. Fitoterapia. 2006;77(5):408-410. doi:10.1016/j.fitote.2006.05.004.
5. Jin X, Ruiz Beguerie J, Sze DM-Y, Chan GCF. Ganoderma lucidum (Reishi mushroom) for cancer treatment. Cochrane database Syst Rev. 2016;4:CD007731. doi:10.1002/14651858.CD007731.pub3.
6. Zhu Q, Bang TH, Ohnuki K, Sawai T, Sawai K, Shimizu K. Inhibition of neuraminidase by Ganoderma triterpenoids and implications for neuraminidase inhibitor design. Sci Rep. 2015;5(AUGUST):13194. doi:10.1038/srep13194.
7. Teplyakova T V., Psurtseva N V., Kosogova TA, Mazurkova NA, Khanin VA, Vlasenko VA. Antiviral Activity of Polyporoid Mushrooms (Higher Basidiomycetes) from Altai Mountains (Russia). Int J Med Mushrooms. 2012;14(1):37-45. doi:10.1615/IntJMedMushr.v14.i1.40.
8. Gao L, Sun Y, Si J, et al. Cryptoporus volvatus extract inhibits influenza virus replication in vitro and in vivo. PLoS One. 2014;9(12). doi:10.1371/journal.pone.0113604.
9. Van Thu Nguyen, Nguyen The Tung, To Dao Cuong, Tran Manh Hung, Jeong Ah Kim, Mi Hee Woo, Jae Sue Choi, Jeong-Hyung Lee, Byung Sun Min, Cytotoxic and anti-angiogenic effects of lanostane triterpenoids from Ganoderma lucidum, Phytochemistry Letters, Volume 12, 2015, Pages 69-74, ISSN 1874-3900,
10. Bhardwaj N, Katyal P, Sharma AK. Suppression of inflammatory and allergic responses by pharmacologically potent fungus Ganoderma lucidum. Recent Pat Inflamm Allergy Drug Discov. 2014;8(2):104-17. doi: 10.2174/1872213×08666140619110657. PMID: 24948193.
11. Liu C, Cao M, Yang N, Reid-Adam J, Tversky J, Zhan J, Li XM. Time-dependent dual beneficial modulation of interferon-γ, interleukin 5, and Treg cytokines in asthma patient peripheral blood mononuclear cells by ganoderic acid B. Phytother Res. 2022 Mar;36(3):1231-1240. doi: 10.1002/ptr.7266. Epub 2022 Feb 3. PMID: 35112740.
12. Friedman M. Chemistry, Nutrition, and Health-Promoting Properties of Hericium erinaceus (Lion’s Mane) Mushroom Fruiting Bodies and Mycelia and Their Bioactive Compounds. J Agric Food Chem. 2015;63(32):7108-7123. doi:10.1021/acs.jafc.5b02914
13. Teekachunhatean, S., Sadja, S., Ampasavate, C., Chiranthanut, N., Rojanasthien, N., & Sangdee, C. (2012). Pharmacokinetics of Ganoderic Acids A and F after Oral Administration of Ling Zhi Preparation in Healthy Male Volunteers. Evidence-Based Complementary and Alternative Medicine, 2012, 780892. https://doi.org/10.1155/2012/780892
14. Verhoeven J, Keller D, Verbruggen S, Abboud KY, Venema K. A blend of 3 mushrooms dose-dependently increases butyrate production by the gut microbiota. Benef Microbes. 2021;12(6):601-612. doi:10.3920/BM2021.0015
Polysaccharides are a major component of the fungal cell wall and possibly the most potent and abundant therapeutic constituent in mushroom medicine. They are well-touted for their immune regulatory activity, but they play a role in many other important physiologic processes including blood sugar regulation, inflammation modulation, antioxidant activity, mood support, and modulation of the intestinal microbiome and the greater intestinal environment.
Biologic Response Modifiers
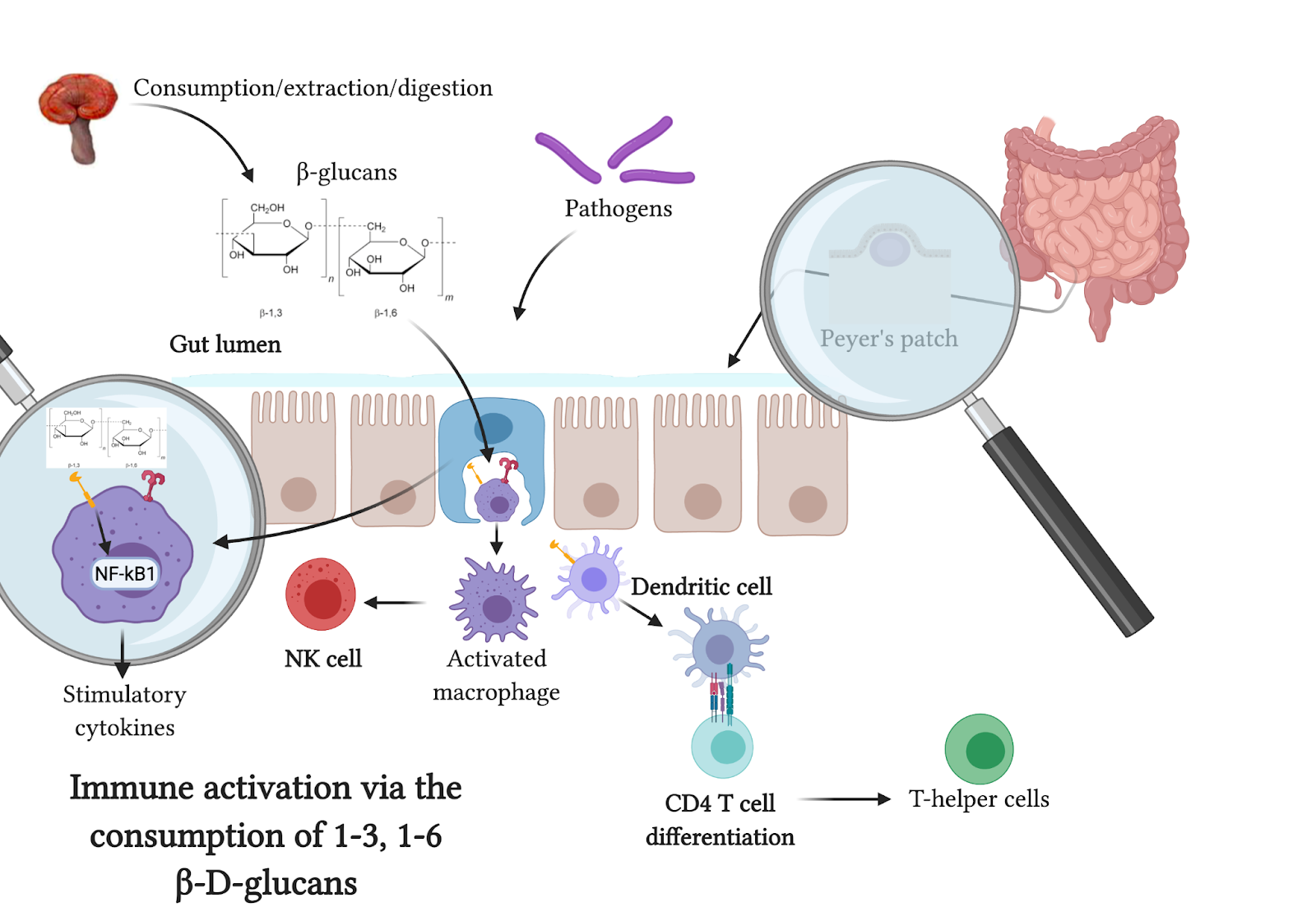
The fungal cell wall is composed of many layers – mannoproteins, ß- (1,6)-glucan and ß-(1,3)-glucan, and chitin. Humans do not make chitinase, and cannot utilize chitin for anything other than non-digestible fiber. Chitin can, however, be broken down by different processing techniques, including heat, allowing the mannan and beta-glucan layers to be utilized as medicine. From an immunological perspective, these layers are considered pathogen-associated molecular patterns (PAMPs), and bind to pathogen recognition receptors (PRRs) on cells in the innate immune system – macrophages, granulocytes, dendritic cells and natural killer cells. The majority of binding occurs in the gut-associated lymphoid tissue (GALT), where Peyer’s patches contain cells of the innate immune system. Specific receptors that bind these ß-glucans include Dectin-1 and TLR-2 (1). Binding has a hormetic effect, creating a stress response in our immune system, stimulating the release of cytokines with anti-inflammatory, pro-inflammatory, and other immune-modulating actions. These cytokines stimulate an increase in natural killer cells and cytotoxic T cells, generating a stress signal that leads to a more responsive immune system.
Blood Sugar regulation
ß-glucan polysaccharides from mushrooms inhibit the enzyme alpha-glucosidase, which is essential for carbohydrate metabolism and for generating a rise in blood sugar following a meal. They have also been shown to decrease gastric emptying, slowing the absorption of glucose, and preventing hyperglycemic episodes. Finally, polysaccharides demonstrate the ability to upregulate GLUT 4, an insulin-response glucose transporter, while down-regulating NF-κB, a nuclear transcription factor that regulates inflammation (2,3).
Antioxidant
ß-glucan polysaccharides increase the activity of hepatic oxidative enzymes such as catalase, glutathione peroxidase, and superoxide dismutase, increasing levels of glutathione and decreasing malondialdehyde levels (a marker of lipid peroxidation). Generally, these compounds support our innate antioxidant systems, relieving oxidative stress, and reducing free radical damage (4).
Prebiotic
The oligosaccharides and polysaccharides contained in mushrooms are consumed by Bifidobacterium and Lactobacilli, two important bacterial phyla in a healthy microbiome (5,6). Polysaccharides also promote production of important postbiotics like butyric acid, a short chain fatty acid, which provides colon cells with about 70% of their total energy needs (7).
Psychobiotic
Polysaccharides play a role in creating a healthy intestinal ecosystem through potentiating the growth of healthy bacteria, archaea and fungi while also supporting a more resilient gastrointestinal terrain. Healthy gut flora and intestinal mucosa are associated with better mood. Signals from the GI tract can independently relay information to the central nervous system, and the maintenance of a healthy GI tract supports a balanced central nervous system. There are a number of studies that have demonstrated increased probiotic intake associated with increased mood and diminished anxiety (8). Specifically, probiotic supplementation with Lactobacillus helveticus and Bifidobacterium longum showed less self-reported negative mood and decreased urinary cortisol. A similar effect was also observed in healthy participants who consumed a mixture of Bifidobacterium bifidum and Bifidobacterium lactis, and Lactobacillus acidophilus, Brevibacillus brevis, Brevibacterium casei, Bifidobacterium salivarius, and Lactococcus lactis (9,10).
Absorption
Raw mushroom preparations (including products labeled “mushroom powder”) have not undergone sufficient processing to break down the chitin in the cell wall, leaving ß-glucans unavailable for absorption; extractions employ heat, water, and alcohol to free ß-glucans, improving their absorption and activity. While mixing mushrooms with coffee, tea, or chocolate is increasingly popular, the tannins in these foods bind the fungal polysaccharides, diminishing bioavailability (11). For best results, take polysaccharide-rich mushroom preparation on an empty stomach, away from food. To learn more, check out my blog post on factors impacting the absorption of mushroom constituents.
DIY Extraction of Polysaccharides
Polysaccharides are water soluble and high molecular weight (heavy molecules that don’t evaporate)
Boil chopped (fresh or dried) mushrooms in water for about 2 hours. Use this water as a base for creams, teas, stews, soups and concentrated extracts
References
Baert K, Sonck E, Goddeeris BM, Devriendt B, Cox E. Cell type-specific differences in β glucan recognition and signalling in porcine innate immune cells. Dev Comp Immunol. 2015;48(1):192-203. doi:10.1016/j.dci.2014.10.005.
Vitak T, Yurkiv B, Wasser S, Nevo E, Sybirna N. Effect of medicinal mushrooms on blood cells under conditions of diabetes mellitus. 2017;8(5):187-201. doi:10.4239/wjd.v8.i5.187.
Chen Y, Liu Y, Rahman M, Yan X, Yang C. Structural characterization and antidiabetic potential of a novel heteropolysaccharide from Grifola frondosa via IRS1 / PI3K-JNK signaling pathways. Carbohydr Polym. 2018;198(15):452-461. doi:10.1016/j.carbpol.2018.06.077.
Wang H, Liu YM, Qi ZM, et al. An Overview on Natural Polysaccharides with Antioxidant Properties. 2015;(February). doi:10.2174/0929867311320230006.
Yu Z, Liu B, Mukherjee P, Newburg DS. Trametes versicolor Extract Modifies Human Fecal Microbiota Composition In vitro. 2013:107-112. doi:10.1007/s11130-013-0342-4.
Palacios S, Losa F, Dexeus D, Cortés J. Beneficial effects of a Coriolus versicolor – based vaginal gel on cervical epithelization , vaginal microbiota and vaginal health : a pilot study in asymptomatic women. 2017:4-9. doi:10.1186/s12905-017-0374-2.
Verhoeven J, Keller D, Verbruggen S, Abboud KY, Venema K. A blend of 3 mushrooms dose-dependently increases butyrate production by the gut microbiota. Benef Microbes. 2021 Nov 16;12(6):601-612. doi: 10.3920/BM2021.0015. Epub 2021 Sep 30. PMID: 34590532.
Messaoudi M, Violle N, Bisson J-F, Desor D, Javelot H, Rougeot C. Beneficial psychological effects of a probiotic formulation (Lactobacillus helveticus R0052 and Bifidobacterium longum R0175) in healthy human volunteers. Gut Microbes 2011;2:256– 261.
Steenbergen L, Sellaro R, van Hemert S, Bosch JA, Colzato LS. A randomized controlled trial to test the effect of multispecies probiotics on cognitive reactivity to sad mood. Brain Behav Immun 2015;48:258–264.
Allen AP, HutchW, Borre YE, et al. Bifidobacterium longum 1714 as a translational psychobiotic: modulation of stress, electrophysiology and neurocognition in healthy volunteers. Transl Psychiatry 2016;6:e939.
Li R, Zeng Z, Fu G, Wan Y, Liu C, McClements DJ. Formation and characterization of tannic acid/beta-glucan complexes: Influence of pH, ionic strength, and temperature. Food Res Int. 2019;120:748-755. doi:10.1016/j.foodres.2018.11.034