Therapeutic dosing: Higher doses, often more frequently throughout the day
Maintenance: Lower doses, often less frequently throughout the day
Immune-Specific Dosing
The concept of therapeutic and maintenance dosing is based on the theory of ‘trained immunity’ (1,2). This theory suggests that ongoing, high doses of ß-glucan-rich mushrooms are not, in fact, necessary to potentiate immune modulation and protection from infections. The innate immune system, or the “first responders,” has the ability to create non-specific memory responses to a variety of subsequent infections after a primary infection or vaccination. For example, a person may become infected with a specific pathogen (virus, bacteria or fungi) and the innate immune system will mount a response to that specific pathogen. Additionally, innate memory cells will remain that can help protect against that same infection in the future, as well as other unrelated infections, i.e. a non-specific memory response. These changes in immune memory have been observed for as long as one year following initial administration of ß-glucans (3). These innate memory cells have a relatively short lifespan, and so it seems unlikely that trained immunity would last as long as it does. However, researchers have discovered that ß-glucans not only induce trained immunity in mature innate immune cells, but they also alter the cells hematopoietic progenitors (cells that will eventually differentiate into innate immune cells) within the bone marrow leading to the prolonged presence of trained monocytes and macrophages in circulation (4). The ß-glucans are essentially playing a role in your immune cells’ epigenetic purpose.
Initial higher dosing of ß-glucan-rich mushrooms is like an immune training program. Similar to building muscle, the initial training takes a little longer, and you have to train more often. Maintaining muscle is much easier than building muscle, and sometimes once a week training is all that is needed for muscle maintenance. Similarly, taking mushrooms one to two times per day, and maybe even a few times per week, is all that is needed for immune maintenance.
Initial dosing during the therapeutic phase for immune modulating effects should be done repeatedly, ideally three times per day. One study comparing the duration of the effect of oral dosing versus intravenous dosing of ß-glucans found that the repeated oral dosing “clearly resulted in stronger and longer action stimulation (5).” Intravenous dosing of ß-glucans is mostly available in clinical trials outside of the United States and is not yet common practice in the U.S. Therefore, repeated oral dosing two to three times per day is most likely to optimize initial immune benefits.
Neurocognitive-specific Dosing
When dosing mushrooms like lion’s mane, tremella and oyster mushroom for neurocognitive benefits, the idea of immune training or cellular training does not apply. In the majority of studies exploring the neurocognitive benefits of lion’s mane, researchers have found that many benefits relating to memory and mood were no longer present after discontinuation of supplementation. It is important to note, however, that certain patterns related to mood may be a result of “negative neuroplasticity.” Negative neuroplasticity is the idea that getting stuck in a particular maladaptive pattern, like insomnia, can create neural patterns that are then primed for insomnia. Changing certain behaviors or taking certain medications and/or supplements can help to rewire negative patterns. Mushrooms with neurological actions, such as BDNF-mediated neurogenesis, may help to rewire negative plasticity, and therefore may not be needed on a daily basis as previously thought. While this may be true for mood-related disorders, it is unlikely to be the case for memory and cognitive enhancement. Either way, once daily dosing seems adequate for neurocognitive effects.
Absorption
Mushrooms are composed of many different constituents. These constituents have different physiological activities that are fully dependent on whether they are absorbed, or not absorbed, into the bloodstream from the intestines.
Immune considerations
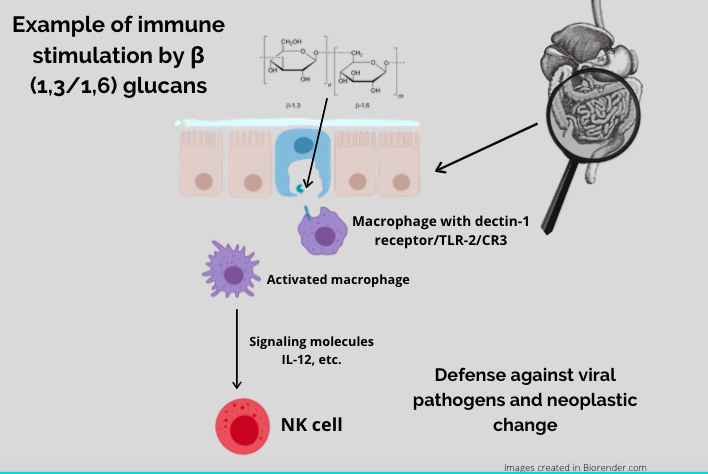
The immune-modulating benefits of mushrooms are dependent on a healthy environment within the small intestine, where there are groups of immune cells within the gut-associated lymphoid tissue (GALT). The GALT contains pockets of immune cells called Peyer’s patches which contain M cells, and M cells present ß-glucans to innate immune cells like macrophages and dendritic cells via specific cell-surface receptors, most notably dectin-1, but also TLR2 and CR3. Mushroom ß-glucans bind to dectin-1 and stimulate an immune response throughout the bloodstream, lymphatic system, spleen, bone marrow, and lymph nodes.
It is important to note, however, that not all ß-glucans are absorbed into the bloodstream, and that some make their way to the large intestine. Mushroom ß-glucans are prebiotics, acting as food for healthy gut microbes and improving the intestinal microenvironment (6). A healthy intestinal microenvironment is fundamental to human health and research has shown that microbial fermentation of fungal ß-glucans can increase important post-biotic short chain fatty acids like butyrate (7). Improving the intestinal microenvironment does not result in trained immunity, like small intestinal absorption of ß-glucans does, so for prebiotic benefits, daily dosing seems best.
Additional Considerations
Food. If there is a lot of food in the small intestine, there will be less surface area for the ß-glucans to come into contact with M cells, and the immune response will likely be less significant than it would be on an empty stomach.
Alcohol. Chronic alcohol consumption has been shown to alter the innate immune system at the level of the Peyer’s patch (8). Frequent alcohol intake may lessen the immune benefits of mushroom ß glucans.
Tannins/polyphenols. Blending and emulsifying ß-glucans with tannin-rich substances like coffee, tea and chocolate create a tannin-polysaccharide complex. However, the bonds between polysaccharides and polyphenols are hydrogen bonds and are easily impacted by temperature and pH (acidity). Bonds can be broken at high temperatures between 68°F-104°F, so if blending mushroom powders with tannin-rich foods, blend at higher temperatures rather than cold. If polysaccharides are contained in these complexes when they reach the small intestine, they will not be available to bind immune receptors in the GALT and their immune-modulating properties will likely be nullified (9).
Work Cited
Moerings, B. G. J., de Graaff, P., Furber, M., Witkamp, R. F., Debets, R., Mes, J. J., van Bergenhenegouwen, J., & Govers, C. (2021). Continuous Exposure to Non-Soluble β-Glucans Induces Trained Immunity in M-CSF-Differentiated Macrophages. Frontiers in Immunology, 12. https://doi.org/10.3389/fimmu.2021.672796
Garcia-Valtanen P, Guzman-Genuino RM, Williams DL, Hayball JD, Diener KR. Evaluation of trained immunity by β-1, 3 (d)-glucan on murine monocytes in vitro and duration of response in vivo. Immunol Cell Biol. 2017 Aug;95(7):601-610. doi: 10.1038/icb.2017.13. Epub 2017 Feb 23. PMID: 28228641; PMCID: PMC5550561.
Moorlag SJCFM, Khan N, Novakovic B, Kaufmann E, Jansen T, van Crevel R, Divangahi M, Netea MG. β-Glucan Induces Protective Trained Immunity against Mycobacterium tuberculosis Infection: A Key Role for IL-1. Cell Rep. 2020 May 19;31(7):107634. doi: 10.1016/j.celrep.2020.107634. PMID: 32433977; PMCID: PMC7242907.
Mitroulis I., Ruppova K., Wang B., Chen L.S., Grzybek M., Grinenko T., Eugster A., Troullinaki M., Palladini A., Kourtzelis I. Modulation of Myelopoiesis Progenitors Is an Integral Component of Trained Immunity. Cell. 2018;172:147–161.e12.
Vetvicka, Vaclav & Vetvickova, Jana & Ms,. (2008). A Comparison of Injected and Orally Administered β-glucans. 11.
Nowacka-Jechalke N, Juda M, Malm A. The preliminary study of prebiotic potential of Polish wild mushroom polysaccharides: the stimulation effect on Lactobacillus strains growth. Eur J Nutr. 2018 Jun;57(4):1511-1521. doi: 10.1007/s00394-017-1436-9. Epub 2017 Mar 28. PMID: 28353071; PMCID: PMC5959981.
Mitsou EK, Saxami G, Stamoulou E, Kerezoudi E, Terzi E, Koutrotsios G, Bekiaris G, Zervakis GI, Mountzouris KC, Pletsa V, Kyriacou A. Effects of Rich in Β-Glucans Edible Mushrooms on Aging Gut Microbiota Characteristics: An In Vitro Study. Molecules. 2020 Jun 18;25(12):2806. doi: 10.3390/molecules25122806. PMID: 32570735; PMCID: PMC7355846.
Li R, Zeng Z, Fu G, Wan Y, Liu C, McClements DJ. Formation and characterization of tannic acid/beta-glucan complexes: Influence of pH, ionic strength, and temperature. Food Res Int. 2019;120:748-755. doi:10.1016/j.foodres.2018.11.034
Proving that mushrooms are an essential part of the human diet
Definitions to consider while reading:
Antioxidant: Compounds that mitigate oxidation. Oxidation is a reaction that can produce free radicals, leading to cellular damage. Antioxidants such as thiols and ascorbic acid (vitamin C) terminate these reactions by donating electrons. To balance oxidative states, living organisms maintain systems with overlapping antioxidants such as glutathione, catalase, superoxide dismutase.
Blood-brain barrier: a filtering mechanism of the capillaries that carry blood to the brain and spinal cord tissue, blocking the passage of certain substances.
Cation chelator: a type of bonding of ions and molecules to metal ions. There is application of this in removing toxic metals from the body.
Glutathione: Glutathione is a tripeptide (cysteine, glycine, and glutamic acid) found in surprisingly high levels—5 millimolar—concentrations in most cells. Responsible for the following actions in the body: (Pizzorno)
Direct chemical neutralization of singlet oxygen, hydroxyl radicals, and superoxide radicals
Cofactor for several antioxidant enzymes
Regeneration of vitamins C and E
Neutralization of free radicals produced by Phase I liver metabolism of chemical toxins
One of approximately 7 liver Phase II reactions, which conjugate the activated intermediates produced by Phase I to make them water soluble for excretion by the kidneys
Transportation of mercury out of cells and the brain
Regulation of cellular proliferation and apoptosis
Vital to mitochondrial function and maintenance of mitochondrial DNA (mtDNA)
Lipid peroxidation: The oxidative degradation of lipids. It is the process in which free radicals “steal” electrons from the lipids in cell membranes, resulting in cell damage. (Wik)
Oxidative stress: disturbance in the balance between the production of reactive oxygen species (free radicals) and antioxidant defenses (Betteridge)
Pleurotus ostreatus – Oyster Mushroom
‘Constituentism’
I am not necessarily a fan of isolation and ‘constituentism’ in supplemental medicine – especially when a constituent has been isolated from its synergistic community of molecules from the whole plant or whole mushroom and is considered supposedly different than a pharmaceutical. Though, I do love to understand the individual constituents in organisms and how they work independently, in order to better understand how they work synergistically. There is also a greater understanding to be had of how animals, plants, fungi and bacteria all evolved together when we can explore specific plant and fungi constituent mechanisms with specific cellular receptors and transporters in the animal organism. A specific example of this is a compound called Ergothioneine (ERG). ERG is an amino acid derivative, specifically a crystalline betaine, derivative of histidine – the etymology explains; ergot- (found in ergot (fungus) thione- (double bond to a Sulphur) and -ine (amine containing)—basically a sulphur containing amine found in fungi. ERG is also found in Actinobacteria (gram positive bacteria that behave similar to fungi in soil, helping to decompose the organic matter of dead organisms so the molecules can be taken up by new plants, they even grow extensive mycelium like fungi and were in fact long believed to be fungi) and Cyanobacteria, commonly known as blue-green algea, (another abnormal bacteria that can actually obtain their energy from photosynthesis – they are the only photosynthetic prokaryote able to produce oxygen). (blankenfeldt)
Specific ERG transporters in the animal
We have evolved so closely with mushrooms that we have specific transporters and receptors uniquely for mushroom compounds. ERG is no exception. While we have a specific receptor for ERG, we cannot synthesize this molecule ourselves, and as I will describe later, we are discovering this compound is extremely important for human health. The ERG transporter, OCTN1, (gene: SLC22A4) is found specifically on erythrocytes, fetal liver and bone marrow, ileum of the small intestine, trachea, kidney, cerebellum, lung, monocytes, seminal vesicles and the lens and cornea of the eye (Halliwell). Interestingly, ERG transporter is concentrated in the mitochondria of cells – suggesting a role in protecting mitochondrial components from DNA damage (Paul, Halliwell). This transporter’s only known role is to sequester as much ERG within the cells as is available, and only cells with this transporter can absorb, distribute, and retain this compound (Grundermann). Researchers explored the effects when cells were depleted of this transporter, the results being that cells were more susceptible to oxidative stress, leading to mitochondrial damage, protein oxidation, and lipid peroxidation. Once taken up into the cell, ERG is extremely bioavailable and is retained for up to a month within the body.
Bioavailability of ergothioneine from mushroom consumption
Once consumed in the diet, whether it be the isolated molecule or from whole mushrooms, ERG is quickly absorbed into the blood stream. Human erythrocytes (red blood cells, RBC) contain 2-9 fold more ERG than plasma. Interestingly, concentration is lowest in early life (1-10yo), increases between 11-18yo and reaches maximum value of 3.7mg/100mL by age 18. In this study, subjects consumed 8g and 16g of mushrooms and increases in red blood cell ERG were observed. After 1 and 4hr of consumption, the 16g mushroom dose increased RBC ERG concentration compared to the control, and after only 2 hrs of 16g of mushroom consumption, RBC ERG concentration was significantly higher than the control. Variability in bioavailablity was due to genetic variations in the SLC22A4 gene (Heller, Kris-etherton, Beelman). Interestingly, mutations in the ERG transporter have been identified as a susceptibility factor for autoimmune disorders, such as rheumatoid arthritis and Crohn’s disease, as well as neurodegenerative disorders, such as dementia and Parkinson’s (Paul, Snyder).
Why do we care – Once ergothioneine is in the cell, then what happens?
Ergothioneine as an antioxidant and cytoprotectant: The distribution of ERG transporters may seem random, but on closer inspection, they seem to be present in tissues predisposed to high levels of oxidative stress and inflammation (Halliwell). In animal studies, animals who were completely deficient in ERG had higher levels of reactive oxygen species and were therefore more susceptible to oxidative stress. In vitro, ERG is a powerful scavenger of hydroxyl radical and has been shown to deactivate singlet oxygen at a higher rate than glutathione (Hseu, Servillo). Similar results were proven in vivo – rats supplemented with ERG had lower levels of lipid peroxidation and higher levels of glutathione and alpha-tocopherol. As one might surmise, ERG and glutathione seem to have an intimate relationship within the cell. Glutathione is considered the major intracellular antioxidant in almost all organisms and has important functions in detoxification and immune function. It has been proposed, based on current research, that ERG can help maintain GSH levels in the presence of oxidative burden by interacting with other cellular defense systems. The maintenance of glutathione tissue levels is important in maintaining health as depletion will impair immune function. Conveniently, mushrooms contain both glutathione and ERG. In fact, mushrooms have been observed to have higher GSH amounts than any vegetable or fruit. Grifola frondosa (Maitake), Hericium erinaceus (Lions mane), Pleurotus ostreatus (Oyster mushroom), Boletus edulis (porcini) and Lentinus edodes (shitake) containing the most, respectively (Kalaras). Considering the relationship between ERG and GSH in mushrooms, a correlation analysis found that mushrooms high in GSH were also high in ERG, specifically the caps or pileus of the mushrooms (kalaras). All this said, mushrooms are an important source for cellular antioxidants.
Cation chelator: ERG chelates divalent metal cations – specifically, Cu2, hg, Zn, Cd, Co, Fe, and Ni (Cheah, Song, Kerley). Binding these cations in the body may help prevent their participation in the generation of reactive oxygen species. For example, EGT has been found to protect DNA and protein against copper induced oxidative damage through formation of a redox-inactive EGT-Cu complex. Interestingly, the high levels of ERG in semen – due to the high concentration of ERG OCTN1 transporter on the seminal vesicles- have been shown to prevent Cu inhibition of sperm motility. There is more to be explored between the relationship of ERG and semen viability.
Ergothioneine, aging, and cognitive decline: As we age, we are more vulnerable to the oxidative stress and environmental toxins that slowly damage our DNA, making us more susceptible to neurodegeneration – one of many conditions associated with DNA damage from oxidative stress. Low levels of glutathione have been linked to certain neurodegenerative diseases including Parkinson’s disease (Wei, Shah). ERG was able to dose dependently enhance glutathione activity in the rat liver cytosol—leading researchers to postulate that declining ERG may play a role in age related decline of GSH and glutathione peroxidase. Furthermore, ERG levels were found to be lower in the elderly with early stages of dementia and in PD patients relative to age matched healthy controls (Cheah). In animal studies, oral administration of ERG protected neurons and preserved cognitive function following administration of toxic amyloid beta cisplatin or D-galactose. It is now understood that the presence of Octn1 transporter in the blood brain barrier is responsible for these neuroprotective actions. Captivatingly, researchers found that there is a direct relationship between whole blood and brain ERG levels following consumption (Cheah). It is rare for compounds to be bioavailable in this way and transported across the blood brain barrier so readily – even glutathione needs to be taken intranasal for treatment of neurodegeneration.
Ergothioneine human trial: uptake metabolism and effects
While there is undeniably a lack of human trials exploring the in vivo effects of ERG, a recent study from 2017 explored ERG uptake, metabolism, and effects on biomarkers of oxidative damage and inflammation in healthy human subjects (cheah, Tang). One of the most interesting aspects of ERG discussed in this particular article is that ERG is a tautomer. This means that it exists in two forms – thione and thiol. The thiol is a single bond to sulfur and the thione is a double bond to sulfur. The article explains that in animal physiologic conditions, ERG primarily exists as the thione tautomer – under circumstances of low stress in the body, ERG remains in its thione tautomer form and is not the first choice as an antioxidant, rather endogenous antioxidants like glutathione are preferred (glutathione is a primary antioxidant thiol in the body). When the cells undergo higher levels of oxidative stress, ERG transforms into its thiol tautomer form and is then used for extra support. Additionally, under levels of elevated stress, tissues have increased amounts of the ERG, supposedly by upregulating expression of the OCTN1 ERG transporter in response to inflammatory cytokines.
This study also found that ERG can be stored in the cells for up to a month. It is theorized that ERG is stored for an extended period of time following consumption until it is required as a stronger defense mechanism.
During administration of ERG, plasma levels significantly elevated, while whole blood levels steadily increased for up to four weeks after administration stopped (red blood cells could continue to take ERG up as needed), and the excretion of ERG in the urine remained low, indicating that ERG is absorbed and retained in the body after oral administration. While this study used pure ERG, other studies previously mentioned (Heller) prove this same high bioavailability of ERG from dietary mushrooms, and so I will consider this as true for dietary ERG.
Mushroom analyses of ergothioneine: Fruiting body or mycelium? Gilled mushrooms or polypore mushrooms?
ERG was highest in the fruiting body of Pleurotus ostreatus and in the mycelium of Pleurotus eryngii. Generally, fruiting bodies and mycelium contained different amounts of ERG, with Pleurotus genus containing the highest amounts overall (Chen). Among simple mushrooms, fruiting body of White Button had the least (1.4mg erg per 85g mushroom) and Portabella the highest (2.7mg erg per 85g mushroom). While among specialty mushrooms, Maitake had the least (16.3mg erg per 85g mushroom) and oyster the highest (26.4mg per 85g mushroom) (Dubost).
Fruiting bodies of gilled mushrooms, specifically the cap, contain the most ERG, while polypore mushrooms contain very small amounts. Interestingly, the mycelium of Ganoderma contains more ERG than the fruiting body, but still not as much as the fruiting bodies of Shitake, Matsutake, Oyster, and Maitake (Lee, Wang).
Extraction:
Hot water extraction is sufficient to extract ERG from mushrooms.
In a study that explored water extraction – temperature of water, ratio of water to mycelium and the extraction rate of ERG, the following information was found:
Most notable: at 85°C and 20:1 ratio of mycelium to water, the extraction rate of ERG was 91.2% – the least effective yield of ERG was at 78°C and a 20:1 ratio of mycelium to water. There was no difference in ERG concentration between 5 minutes and 120 minutes, so a long boil to extract EGR is not necessary (Zhang).
In Summary:
Ergothioneine is a water soluble compound that is most abundant in Oyster mushrooms. There are transporters on different tissues in the body that are highly specific to ergothioneine. Ergothioneine is readily absorbed into the blood after consumption of mushrooms and stored in tissues for up to 1 month. In times of excessive oxidative stress, ergothioneine is taken up by those tissues and used as an antioxidant. Of note: there are transporters on the blood brain barrier and there is an association with low ergothioneine and age-related cognitive decline. In a world full of environmental toxins that are mostly impossible to escape, we might as well eat more mushrooms and get some extra protection.
How to apply this information to your life:
Eat a lot of mushrooms, especially Oyster mushrooms.
Antonicelli F, Aruoma OI. Ergothioneine inhibits oxidative stress- and TNF- a -induced NF- j B activation and interleukin-8 release in alveolar epithelial cells. 2003;302:860-864. doi:10.1016/S0006-291X(03)00224-9.
Aruoma OI, Spencer JPE, Mahmood N. Protection Against Oxidative Damage and Cell Death by the Natural Antioxidant Ergothioneine. 1999;37.
Aruoma OI, Whiteman M, England TG, Halliwell B. Antioxidant Action of Ergothioneine : Assessment of Its Ability to Scavenge Peroxynitrite. 1997;391(231):389-391.
Aubert DIRKT. Dietary Sources and Antioxidant Effects of Ergothioneine. 2007:6466-6474. doi:10.1021/jf071328f.
Benson KF, Ager DM, Landes B, Aruoma OI, Jensen GS. Improvement of joint range of motion ( ROM ) and reduction of chronic pain after consumption of an ergothioneine-containing nutritional supplement. Prev Med (Baltim). 2018;54(2012):S83-S89. doi:10.1016/j.ypmed.2012.02.001.
Betteridge, JD What Is Oxidative Stress? 2000:3-8.
Blankenfeldt W, Seebeck FP. Ergothioneine Biosynthetic Methyltransferase EgtD Reveals the Structural Basis of Aromatic Amino Acid Betaine Biosynthesis. 2015:119-125. doi:10.1002/cbic.201402522.
Cheah IK, Halliwell B. Biochimica et Biophysica Acta Ergothioneine ; antioxidant potential , physiological function and role in disease ☆. BBA – Mol Basis Dis. 2012;1822(5):784-793. doi:10.1016/j.bbadis.2011.09.017.
Chen S, Ho K, Hsieh Y, Wang L, Mau J. LWT – Food Science and Technology Contents of lovastatin , g -aminobutyric acid and ergothioneine in mushroom fruiting bodies and mycelia. LWT – Food Sci Technol. 2012;47(2):274-278. doi:10.1016/j.lwt.2012.01.019.
Deiana M, Rosa A, Casu V, et al. modulates oxidative damage in the kidney and liver of rats in vivo : studies upon the profile of polyunsaturated fatty acids L -Ergothioneine. 2004;5614:183-193. doi:10.1016/S0261-5614(03)00108-0.
Dubost NJ, Ou B, Beelman RB. Food Chemistry Quantification of polyphenols and ergothioneine in cultivated mushrooms and correlation to total antioxidant capacity. 2007;105:727-735. doi:10.1016/j.foodchem.2007.01.030.
Franzoni F, Colognato R, Galetta F, et al. An in vitro study on the free radical scavenging capacity of ergothioneine : comparison with reduced glutathione , uric acid and trolox. 2006;60:453-457. doi:10.1016/j.biopha.2006.07.015.
Gruber J, Fong S, Chen C, et al. Mitochondria-targeted antioxidants and metabolic modulators as pharmacological interventions to slow ageing. Biotechnol Adv. 2013;31(5):563-592. doi:10.1016/j.biotechadv.2012.09.005.
Gründemann D. The ergothioneine transporter controls and indicates ergothioneine activity — A review. Prev Med (Baltim). 2018;54(2012):S71-S74. doi:10.1016/j.ypmed.2011.12.001.
Halliwell B, Cheah IK, Drum CL. Biochemical and Biophysical Research Communications Ergothioneine , an adaptive antioxidant for the protection of injured tissues ? A hypothesis. Biochem Biophys Res Commun. 2016;470(2):245-250. doi:10.1016/j.bbrc.2015.12.124.
Harlfinger S, Golz S, Geerts A, et al. Discovery of the ergothioneine transporter. 2005.
Hseu Y, Lo H, Korivi M, Tsai Y, Tang M. Free Radical Biology and Medicine Dermato-protective properties of ergothioneine through induction of Nrf2 / ARE-mediated antioxidant genes in UVA-irradiated Human keratinocytes. Free Radic Biol Med. 2015;86(91):102-117. doi:10.1016/j.freeradbiomed.2015.05.026.
Kalaras MD, Richie JP, Calcagnotto A, Beelman RB. Mushrooms: A rich source of the antioxidants ergothioneine and glutathione. Food Chem. 2017;233:429-433. doi:10.1016/j.foodchem.2017.04.109.
Kerley RN, Mccarthy C, Kell DB, Kenny LC. Free Radical Biology and Medicine The potential therapeutic e ff ects of ergothioneine in pre-eclampsia. 2018;117(August 2017):145-157. doi:10.1016/j.freeradbiomed.2017.12.030.
Lee WY, Park E-J, Ahn JK, Ka K-H. Ergothioneine Contents in Fruiting Bodies and Their Enhancement in Mycelial Cultures by the Addition of Methionine. Mycobiology. 2009;37(1):43. doi:10.4489/MYCO.2009.37.1.043.
Lo Y, Lin S, Ulziijargal E, et al. Comparative Study of Contents of Several Bioactive Components in Fruiting Bodies and Mycelia of Culinary-Medicinal Mushrooms. 2012;14(4):357-363.
Li RWS, Yang C, Sit ASM, et al. Uptake and Protective Effects of Ergothioneine in Human Endothelial Cells. 2014;(September):691-700.
Markova NG, Karaman-jurukovska N, Dong KK, Damaghi N, Smiles KA, Yarosh DB. Free Radical Biology & Medicine Skin cells and tissue are capable of using L -ergothioneine as an integral component of their antioxidant defense system. Free Radic Biol Med. 2009;46(8):1168-1176. doi:10.1016/j.freeradbiomed.2009.01.021.
Martin KR. The Bioactive Agent Ergothioneine, a Key Component of Dietary Mushrooms, Inhibits Monocyte Binding to Endothelial Cells Characteristic of Early Cardiovascular Disease. 2010;13(6):1340-1346.
Moncaster JA, Walsh DT, Gentleman SM, Jen L, Aruoma OI. Ergothioneine treatment protects neurons against N -methyl- d – aspartate excitotoxicity in an in vivo rat retinal model. 2002;328:55-59.
Nakamichi N, Nakayama K, Ishimoto T, Masuo Y. Food-derived hydrophilic antioxidant ergothioneine is distributed to the brain and exerts antidepressant effect in mice. 2016;477:1-10. doi:10.1002/brb3.477.
Nguyen TH, Giri A, Ohshima T. A rapid HPLC post-column reaction analysis for the quantification of ergothioneine in edible mushrooms and in animals fed a diet supplemented with extracts from the processing waste of cultivated mushrooms. Food Chem. 2012;133(2):585-591. doi:10.1016/j.foodchem.2012.01.061.
Onofrio ND, Servillo L, Giovane A, et al. Free Radical Biology and Medicine Ergothioneine oxidation in the protection against high-glucose induced endothelial senescence : Involvement of SIRT1 and SIRT6. Free Radic Biol Med. 2016;96:211-222. doi:10.1016/j.freeradbiomed.2016.04.013.
Paul, BD, Snyder, SH. The Unusual Amino Acid, l-ergothioneine is a Physiologic Cytoprotectant. NIH public access 2010: 17-7. Doi: 10.1038/cdd.2009.163
Sakrak O, Kerem M, Bedirli A, et al. Ergothioneine Modulates Proinflammatory Cytokines and Heat Shock Protein 70 in Mesenteric Ischemia and Reperfusion Injury. 2018;42(2008):36-42. doi:10.1016/j.jss.2007.04.020.
Servillo L, Castaldo D, Casale R, et al. Free Radical Biology and Medicine An uncommon redox behavior sheds light on the cellular antioxidant properties of ergothioneine. Free Radic Biol Med. 2015;79:228-236. doi:10.1016/j.freeradbiomed.2014.11.017.
Shah SP, Duda JE. Dietary modifications in Parkinson ’ s disease : A neuroprotective intervention ? Med Hypotheses. 2018;85(6):1002-1005. doi:10.1016/j.mehy.2015.08.018.
Song T, Chen C, Liao J, Ou H, Tsai M. Ergothioneine protects against neuronal injury induced by cisplatin both in vitro and in vivo. Food Chem Toxicol. 2010;48(12):3492-3499. doi:10.1016/j.fct.2010.09.030.
Tomomi I, Manami K, Hironobu T, Etsuko H, Toshio N, Toshihiko O. Ergothioneine as an Anti-Oxidative / Anti-Inflammatory Component in Several Edible Mushrooms. 2011;17(2):103-110.
Wang X, Zhang J, Wu L, et al. A mini-review of chemical composition and nutritional value of edible wild-grown mushroom from China. Food Chem. 2014;151:279-285. doi:10.1016/j.foodchem.2013.11.062.
Weigand-heller AJ, Kris-etherton PM, Beelman RB. The bioavailability of ergothioneine from mushrooms ( Agaricus bisporus ) and the acute effects on antioxidant capacity and biomarkers of in fl ammation. Prev Med (Baltim). 2018;54(2012):S75-S78. doi:10.1016/j.ypmed.2011.12.028.
Wei Z, Li X, Li X, Liu Q, Cheng Y. Oxidative Stress in Parkinson ’ s Disease : A Systematic Review and. 2018;11(July):1-7. doi:10.3389/fnmol.2018.00236.
Yoshida S, Shime H, Funami K, Takaki H. The Anti-Oxidant Ergothioneine Augments the Immunomodulatory Function of TLR Agonists by Direct Action on Macrophages. 2017:1-15. doi:10.1371/journal.pone.0169360.
Forget about the idea that mushrooms have chemicals that have pharmacological potential. Forget that you want to learn about the mushrooms so that you can ‘use’ them.
Just for a moment, let’s see the mushrooms as their own organism.
Their own organism with all of their chemicals not for the human system, but for the fungal system.
Every constituent that we wish to extract and isolate and identify has an important role within the living mushroom, and a role within the entire ecosystem.
Let’s explore these compounds and their functions within the mushroom before exploring their functions and relations within the human system.
Remember, a human is not their cholesterol, a human is not their pheromones, a human is not their amino acids. A human is a synergistic organism made up of thousands of molecules and cells all working together. It is impossible to understand who a human is from just analyzing one of these molecules.
When we look at the mushroom through the lens of its parts, keep in mind it is the synergism between these parts that makes the organism, not the isolated constituents.
The purpose of this writing is to first investigate major myco-constituents that have been researched for their medicinal value and understand their innate role in the fungal organism. After delving into the mushrooms on a cellular level I’ll discuss how these constituents interact with the human system. We did after all evolve with the mushrooms, we just work so beautifully together. Through understanding the innate role of these compounds and where they are within the mushroom organism, we gain insight into the best ways to extract these compounds to support our own physiology. Specific constituents that will be discussed: Polysaccharides, volatile compounds (Alcohols, monoterpenes, sesquiterpenes, aldehydes, ketones), triterpenes, ergosterol, phenolic compounds, and fatty acids.
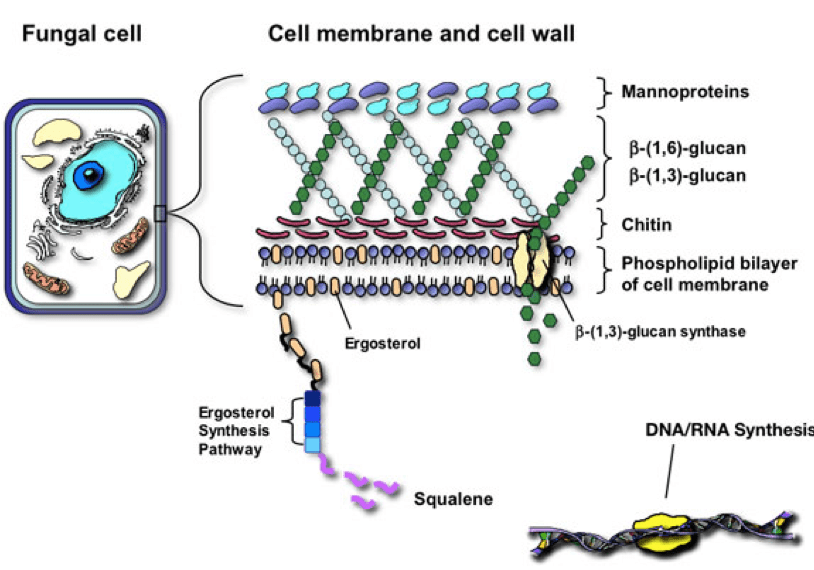
The cell wall is an exquisite and imperative structure in all organisms. The cell wall role in fungi is similar to the role of the cell membrane in humans – it is dynamic in controlling the shape and protecting the organism from the environment. The cell well is made up of compounds involved in morphogenesis, reproduction, cell-cell and cell-matrix interaction. The fungal cell is rigid as it is made up of one of the hardest substances, chitin. Although rigid, it must also be complex to allow for budding, growth and adaptation to environmental stress. The fungal cell wall is composed of multidimensional, communicating polysaccharides including chitin and glucans. (Taborda)
Polysaccharides
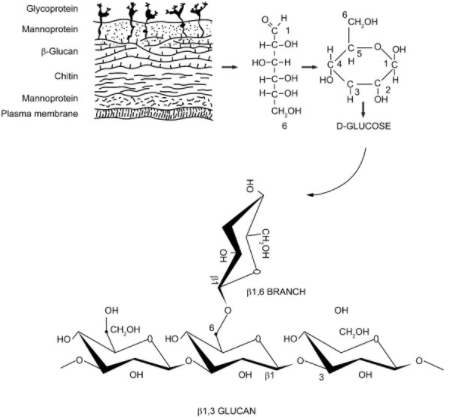
– Many simple sugar molecules attached to one another, referred to as glucans when it is glucose molecules-
These chains of sugars may be extracellular (outside the cell wall), may be associated with the cell wall and membrane, or may be intracellular. Extracellular polysaccharide function is unclear, pullulan a polysaccharide similar to glycogen and mycodextran are alpha linked glucans that are extracellular. From what I could gather, the function of these may be as storage reserves. Beta 1,3 and beta 1,6 glucans are also sometimes found on the outside of the cell, but are most often important components of the cell wall itself. (Griffin) The major role of polysaccharides is in providing cell wall structure. As Basidiomycota (fungi with fruiting bodies) grow and mature fruiting bodies, the total polysaccharide levels increase. (Friedman)
With this knowledge about the role of these compounds as structural support and barrier to the fungal cell we can understand the role within the human organism.
In the Human
The polysaccharides act in the human in an analogous way as they act in the fungal cell. They support the human cells in responding to environmental stressors within the human terrain.
Polysaccharides play a number of functions in the human system. Once ingested, these compounds move through the GI, coming into contact to mucous membranes all the way down through the inner tube from mouth to anus. The mucous membranes have lymphoid tissue called gut associated lymphoid tissue (GALT) that contains immune cells called macrophages. These macrophages, or big eaters, are an important part of our innate immune system. These immune cells have specific receptors for beta-glucans, Dectin-1 and TLR2. Once the beta- glucans come into contact with them, there are chemical messengers called cytokines that are stimulated. Specifically, IL-12 and IFN-y. These cytokines are important in stimulating Natural killer cells, Cytotoxic T cells and TH1 cells. Once these cells are stimulated they target viruses and cancer cells. (guggenhein)
Polysaccharides also function as soluble fiber. When they enter the gut, they slow the release of glucose into the blood stream, acting as hypoglycemic compounds. They also help to decrease cholesterol through binding bile acids. When bile acids end up in the soluble fiber, they are pooped out and then the body will use cholesterol to make more, rather than recycling the bile acids. Along with binding bile acids, they also prevent absorption of cholesterol from food in the GI, further lowering serum cholesterol. These polysaccharides are also beneficial in controlling blood sugar. Polysaccharides inhibit the enzyme, alpha-glucosidase, this enzyme is known to induce after meal hyperglycemia. Along with inhibiting this enzyme, polysaccharides have also been shown to upregulate the GLUT 4, insulin-response glucose transporter, while down regulating NFkappa-B, a nuclear transcription factor that controls regulation of inflammation. (friedman, shamtsyan)
Polysaccharides as antioxidants: Polysaccharides increase activities of liver oxidative enzymes, catalase, glutathione peroxidase, and superoxide dismutase and increase glutathione and malondialdehyde levels. This is all to say that these compounds support our innate antioxidant systems supporting the human in relieving oxidative stress and free radical damage.
Polysaccharides as prebiotics: The oligosaccharides and polysaccharides contained in mushrooms are consumed by Bifidobacterium and Lactobacilli, two important bacteria in a healthy microbiome. Through supporting a healthy gut microbiome, this can also be helpful with asthma, allergies, anxiety and even depression.
Extraction method:
Hot water – 100 degrees C for at least 2 hours
Volatiles Compounds and Triterpenes
Low molecular weight, carbon based compounds that vaporize at about 20 degrees Celsius. Include: Alcohols, monoterpenes, sesquiterpenes, aldehydes, and ketones
Triterpenes have 30 carbons and are too heavy to be volatile, but I put them here since they are made in the same pathway as monoterpenes and sesquiterpenes.
These volatile compounds are derived from primary and secondary metabolism pathways; they diffuse through the atmosphere as “infochemicals”
These chemicals function as attractants and deterrents to insects and other invertebrates. Fungal volatiles have important properties as pheromones and defense. Some of these compounds are an efficient way of defending against fungal feeders. Octanol, often called mushroom alcohol, is an alcohol that deters banana slugs from eating mushrooms while also serving as an attractant for fungus eating beetles. Amusingly, octanol from human skin serves as a host odor cue that attracts blood sucking insects. Trametes versicolor is an example of a polypore mushroom that produces sesquiterpenes: specifically, cardinene, beta guiaene, isoledene and gamma patchoulene that attract fungivorous beetles. (morath,Rosecke) Through attracting these insects the mushrooms are better able to spread their spores and inoculate nearby substrate. A good way to make more of themselves!
Monoterpenes, sesquiterpenes and triterpenes are all compounds in mushrooms that have a cholesterol backbone. (The chemical structure looks similar to cholesterol)
Triterpenes are the heaviest of the three since they have 30 carbons and the others have 10 and 15 carbons respectively. Within the mushroom cell, triterpenes are converted into various metabolites including sterols, steroids, and saponins. They are produced via a pathway called the mevalonate pathway with a starting molecule called acetyl-coa. Animals have a very similar pathway that leads to cholesterol synthesis! This pathway in fungi also leads to ergosterol (a precursor to vitamin D2) which plays the same role that cholesterol plays in the human cell. It is a major component of the plasma cell membrane in fungi, important for membrane permeability.
Cytochrome p450 is important in gene expression and triterpenoid production. There is significantly more CYP450 during the transition from primordia to fruiting bodies, correlating with a higher triterpene content in the mushroom fruiting bodies. – developmental stage of fungal growth plays an important role in the regulation of secondary metabolite genes. (rep NP) (Schmidt-dannert, Schrader)
In the Human
Volatile compounds are very low molecular weight and cross easily through barriers and membranes. It is for this reason that essential oils are so readily absorbed through the skin and mucous membranes. Some volatile compounds can be neurotoxic for this reason, and some can be neuroprotective. There is research now exploring the role of octanol as an antiseizure agent. (Manjarrez-Marmolejo, chang, jahromi) Octanol reduced seizure induction and seizure discharges when it was applied directly to the epileptic focus in the somatosensory cortex. Additionally, it has been reported that administration of octanol significantly reduces the frequency and amplitude of epileptiform spikes, as well as the epileptic behavioral score induced by the administration of penicillin. (Manjarrez).
Some fungal sesquiterpenes have also been shown to inhibit TGF-b, decreasing kidney fibrosis in late stage kidney disease. Triterpenes are major compounds of interest in cancer research. Triterpenes have been shown to be directly cytotoxic to many cancer cell lines. (Rios, Grienke, Bhattarai, Ren, Jin, , Zhu) Triterpenes also have specific antiviral activity, inhibiting an enzyme, neuraminidase, which is important for viruses to be able to leave one cell to enter another. (Teplyakova, Gao) Triterpene compounds have been shown to be very beneficial in treatment of hypersensitivity reactions. Hypersensitivity reactions include allergy, asthma, dermatitis, and rhinitis. Triterpenes inhibit histamine release from mast cells – less histamine will result in less of a reaction. These can also be used in treatment of inflammatory reactions caused by insect stings and bites. (Rios)
Extraction Method
Volatile compounds need to be extracted from fresh mushrooms and will escape into the atmosphere with heat. They are alcohol and fat soluble. Best to do a cold ethanol extraction for volatile compounds. 24 hours is sufficient.
Triterpenes will not volatilize and are also fat and alcohol soluble. There are terpenes within the cell, and so it is best to break the chitin cell wall first with a water decoction, before proceeding with ethanol to extract the triterpenes. A triple extraction method will be provided at the end of this post.
Fatty acids
– Molecules that are long chains of lipid-carboxylic acid found in fats and oils and in cell membranes as a component of phospholipids and glycolipids –
In the Mushroom
Principal lipids in mushrooms are palmitic and linoleic acid. The fruiting body and pileus (cap) contains more linoleic acid and the stipe contains for oleic acid. The linoleic acid plays a role in fungal reproductive mechanisms. Trametes versicolor contains more short chain fatty acids – octanoic, decanoic and lauric acid. (Summer) Fungal membranes contain large quantities of free fatty acids, specifically glycerolipids and acylglycerols. These fatty acids occur in fungi as the major constituents of oil droplets suspended in mycelial and spore cytoplasm. They are also minor constituents of membranes and cell walls. Their primary function is storage material. Phophoglycerides are formed from a combination of fatty acids and are found primarily in the plasma membrane of cellular organelles, where they occur as complexes with proteins.
In the Human
Short chain fatty acids found in Trametes species modifies human fecal microbiota composition, increasing healthy bacteria – Bifidiobacteria and Lactobacillus while decreasing E.coli. (Yu) Short chain fatty acids also lower the intestinal pH, inhibiting growth of microbial pathogens (Plummer)
These fatty acid compounds also have antioxidant activities found to be as effective as alpha-tocopherol (Vit E) while also showing significant antimicrobial and anti-fungal activity. (Younis)
Extraction method:
Fat and alcohol soluble
Phenolic compounds
-The term ‘phenolic’ or ‘polyphenol’ can be defined chemically as a substance which possesses an aromatic ring bearing one or more hydroxy substituents, including functional derivatives (esters, methyl ethers, glycosides etc.)- Commonly known phenolic compounds include: bioflavonoids and proanthocyanidins
In the Mushroom
The phenolic compounds in mushrooms are secondary metabolites derived from intermediates of the shikimic acid pathway, the primary role of which is to provide the essential aromatic amino acids phenylananine, tyrosine and tryptophan. The intermediates of the shikimic acid pathway are precursors of aromatic compounds, including phenolic compounds. The biosynthesis of these compounds has demonstrated that they possess enzymes such as ammonia-lyases that convert phenalynine and tyrosine to cinnamic acids. These compounds represent the building blocks for pigments in mushrooms. The phenolic compound hispolon is an example of a yellow pigment in mushrooms. (Velisek)
In the Human
Phenolic compounds act as powerful antioxidants in the human organism. These compounds help to reduce oxidative stress through their own free radical scavenging activity as well as through stimulating the innate human antioxidant systems. (Geng) Phenolic compounds in mushrooms have also been shown to inhibit alpha-glucosidase in rats and significantly help with diabetic complications caused by this compound. A specific phenolic compound, hispolon, found in Phellinus species has been shown to have analgesic and anti-inflammatory effects as well as inhibits the growth of human cancer cells via the inhibition of the cytokine, TGF-beta. Hispolon also activates caspase, an important enzyme that induces cancer cell death. (Govindappa, Huang, Hong, Chang, Wu, Hsieh)
Extraction Method:
Ethanol or Water depending on how polar the phenolic compound is, best to do a triple extraction so you are sure to get them all.
Triple Extraction
The best way to get all of these compounds in one extract is to do a triple extraction method. The first part of the triple extraction is an overnight ethanol extraction. I use 95% ETOH and poor it over finely chopped/shredded fresh mushroom. Let this sit for 24-48 hours. Press the ETOH from the mushroom material and set aside (ETOH extract #1). Next, place the mushrooms from the original extraction into a crockpot or soup pot, cover with water and simmer for 2-12 hrs (The aqueous extract). Next, place these mushrooms and aqueous extract into a jar and leave the jar 1/3 empty. Fill the last 1/3 of the jar with the ETOH extract #1. Let sit (macerate) for a few weeks, shaking and loving daily. Press out mushrooms (the marc) from ETOH/Aqueous solvent (menstruum). Now you have your triple extraction – containing the volatile compounds, the polysaccharides, the triterpenes, phenolic compounds, and fatty acids.
Work Cited
Bhattarai G, Lee Y-H, Lee N-H, et al. Fomitoside-K from Fomitopsis nigra Induces Apoptosis of Human Oral Squamous Cell Carcinomas (YD-10B) via Mitochondrial Signaling Pathway. Biol Pharm Bull. 2012;35(10):1711-1719. doi:10.1248/bpb.12-00297.
Chang H, Sheu M, Yang C, et al. Analgesic Effects and the Mechanisms of Anti-Inflammation of Hispolon in Mice. 2011;2011. doi:10.1093/ecam/nep027.
Chang WP, Wu JJS, Shyu BC. Thalamic Modulation of Cingulate Seizure Activity Via the Regulation of Gap Junctions in Mice Thalamocingulate Slice. PLoS One. 2013;8(5). doi:10.1371/journal.pone.0062952.
Friedman M. Mushroom Polysaccharides : Chemistry and. 2016. doi:10.3390/foods5040080.
Gao L, Sun Y, Si J, et al. Cryptoporus volvatus extract inhibits influenza virus replication in vitro and in vivo. PLoS One. 2014;9(12). doi:10.1371/journal.pone.0113604.
Geng P, Siu KC, Wang Z, Wu JY. Antifatigue Functions and Mechanisms of Edible and Medicinal Mushrooms. Biomed Res Int. 2017;2017. doi:10.1155/2017/9648496.
Grienke U, Kaserer T, Pfluger F, et al. Accessing biological actions of Ganoderma secondary metabolites by in silico profiling. Phytochemistry. 2015;114:114-124. doi:10.1016/j.phytochem.2014.10.010.
Griffin, David H. (State University Of New York, U. Usa. Fungal Physiology. John Wiley And Sons, 1996.
Guggenheim AG, Wright KM, Zwickey HL. Immune Modulation From Five Major Mushrooms: Application to Integrative Oncology. Integr Med. 2014;13(1):32-44
Hong D, Park MINJU, Jang EUNH, Jung BOM, Kim NAMJ, Kim JHO. Hispolon as an inhibitor of TGF ‑ β ‑ induced epithelial ‑ mesenchymal transition in human epithelial cancer cells by co ‑ regulation of TGF ‑ β ‑ Snail / Twist axis. 2017:4866-4872. doi:10.3892/ol.2017.6789.
Hsieh M, Chien S, Chou Y, Chen C, Chen J, Chen M. Phytomedicine Hispolon from Phellinus linteus possesses mediate caspases activation and induces human nasopharyngeal carcinomas cells apoptosis. Eur J Integr Med. 2014;21(12):1746-1752. doi:10.1016/j.phymed.2014.07.013.
Huang G, Hsieh W, Chang H, Huang S, Lin Y, Kuo Y. r -Glucosidase and Aldose Reductase Inhibitory Activities from the Fruiting Body of Phellinus merrillii. 2011:5702-5706. doi:10.1021/jf2003943.
Jahromi SS, Wentlandt K, Piran S, Carlen PL. Anticonvulsant actions of gap junctional blockers in an in vitro seizure model. J Neurophysiol. 2002;88(4):1893. doi:10.1152/jn.00801.2001.
Jin X, Ruiz Beguerie J, Sze DM-Y, Chan GCF. Ganoderma lucidum (Reishi mushroom) for cancer treatment. Cochrane database Syst Rev. 2016;4:CD007731. doi:10.1002/14651858.CD007731.pub3.
Manjarrez-Marmolejo J F-PJ. Gap Junction Blockers: An Overview of their Effects on Induced Seizures in Animal Models. Curr Neuropharmacol. 2016;14(7):759-771. doi:10.2174/1570159X14666160603115942.
Melappa G, Roshan A, Nithi C, Mohummed TS, – C, Poojari CC. Phytochemical analysis and in vitro antioxidant, antimicrobial, anti-inflammatory and cytotoxicity activities of wood rotting fungi, Trametes ochracea. Pharmacogn J. 2015;7(2):136-146. doi:10.5530/pj.2015.2.8.
Morath SU, Hung R, Bennett JW. Fungal volatile organic compounds: A review with emphasis on their biotechnological potential. Fungal Biol Rev. 2012;26(2-3):73-83. doi:10.1016/j.fbr.2012.07.001.
Plummer NT. Part 2: Treatments for Chronic Gastrointestinal Disease and Gut Dysbiosis. 2015;14(I):25-33.
Ren G, Liu XY, Zhu HK, Yang SZ, Fu CX. Evaluation of cytotoxic activities of some medicinal polypore fungi from China. Fitoterapia. 2006;77(5):408-410. doi:10.1016/j.fitote.2006.05.004.
Rep NP. Traversing the fungal terpenome. 2015;31(10):1449-1473. doi:10.1039/c4np00075g.Traversing.
Ríos JL. Effects of triterpenes on the immune system. J Ethnopharmacol. 2010;128(1):1-14 doi:10.1016/j.jep.2009.12.045.
Rösecke J, Pietsch M, König WA. Volatile constituents of wood-rotting basidiomycetes. Phytochemistry. 2000;54(8):747-750. doi:10.1016/S0031-9422(00)00138-2.
Schmidt-dannert C. Biosynthesis of Terpenoid Natural Products in Fungi. 2015;(November 2014):19-61. doi:10.1007/10.
SCHRADER, JENS. BIOTECHNOLOGY OF ISOPRENOIDS. SPRINGER INTERNATIONAL PU, 2016.
Shamtsyan M, Antontceva E, Panchenko A, Petrischev N. HYPERLIPIDEMIC AND HYPOCHOLESTEROLIC ACTION OF SUBMERGE CULTURED MUSHROOMS.
Sumner JL. The fatty acid composition of basidiomycetes. 2012;8643(1973). doi:10.1080/0028825X.1973.10430293.
Taborda CP, Janeiro R De. GLYCOCONJUGATES AND POLYSACCHARIDES OF FUNGAL CELL WALL AND ACTIVATION. 2008:195-208.
Teng F, Bito T, Takenaka S, Yabuta Y, Watanabe F. Vitamin B 12 [ c ‑ lactone], a Biologically Inactive Corrinoid Compound, Occurs in Cultured and Dried Lion ’ s Mane Mushroom ( Hericium erinaceus ) Fruiting Bodies. 2014. doi:10.1021/jf404463v.
Teplyakova T V., Psurtseva N V., Kosogova TA, Mazurkova NA, Khanin VA, Vlasenko VA. Antiviral Activity of Polyporoid Mushrooms (Higher Basidiomycetes) from Altai Mountains (Russia). Int J Med Mushrooms. 2012;14(1):37-45. doi:10.1615/IntJMedMushr.v14.i1.40.
Velíšek J, Cejpek K. Pigments of Higher Fungi : A Review. 2011;29(2):87-102.
Wu Q, Kang Y, Zhang H, Wang H, Liu Y, Wang J. Biochemical and Biophysical Research Communications The anticancer effects of hispolon on lung cancer cells. Biochem Biophys Res Commun. 2014;453(3):385-391. doi:10.1016/j.bbrc.2014.09.098.
Younis AM, Wu F-S, El Shikh HH. Antimicrobial Activity of Extracts of the Oyster Culinary Medicinal Mushroom Pleurotus ostreatus (Higher Basidiomycetes) and Identification of a New Antimicrobial Compound. Int J Med Mushrooms. 2015;17(6):579-590. doi:10.1615/IntJMedMushrooms.v17.i6.80.
Yu Z, Liu B, Mukherjee P, Newburg DS. Trametes versicolor Extract Modifies Human Fecal Microbiota Composition In vitro. 2013:107-112. doi:10.1007/s11130-013-0342-4.
Zhu Q, Bang TH, Ohnuki K, Sawai T, Sawai K, Shimizu K. Inhibition of neuraminidase by Ganoderma triterpenoids and implications for neuraminidase inhibitor design. Sci Rep. 2015;5(AUGUST):13194. doi:10.1038/srep13194.
Medicinal Fungi complement the human system on a complexity of different levels. The awareness of mushrooms for medicinal use is most present in conversations around the immune system, yet there are further dialogues – in more recent research and in deeper exploration of ethno-mycological studies – that mushrooms support just about every system in the human body, not solely the immune system. This post is going to focus on mushrooms and movement. Specifically, how medicinal mushrooms support the human system through different anti-fatigue mechanisms allowing us to move better longer and with reduced risk of injury.
There are several theories of peripheral fatigue, and not surprisingly, many medicinal mushrooms support the bulk of them. Theories range from metabolic depletion; ATP and creatine-phosphate specifically, lactic acid accumulation, oxidative stress with depletion of endogenous anti-oxidant capabilities, muscle and liver glycogen depletion, tissue damage, and central/psychological factors.
Lactic Acid Accumulation
Let’s first focus on lactic acid accumulation. The accumulation of lactic acid during exercise will inhibit energy metabolism and reduce muscular endurance, resulting in fatigue. (The accumulation of lactate can interfere with nerve impulse and therefore muscle contraction) The theory goes, that if lactic acid accumulation can be controlled, then fatigue wont set in as quickly. Medicinal mushrooms such as Lion’s Mane, Hericium erinaceus, Reishi, Ganoderma spp. and Cordyceps spp.
all enhance the rate of lactic acid clearance during exercise or in the case of Cordyceps, can even inhibit production of lactic acid.1,5,7 The polysaccharides in H. erinaceus had a positive effect on the swimming time of mice, significantly increasing their exercise tolerance in a forced swimming model. Ganoderma spp., Tremella spp. and Cordyceps spp. have been shown to stimulate the enzyme lactic acid dehydrogenase (LDH). The increase in LDH activity helps to increase ATP for exercise under anaerobic conditions as well as accelerate removal of lactic acid. From this information, we can postulate that when we have these mushroom extracts in our systems during exercise, we may benefit from a decrease in lactic acid accumulation, and therefore improve our endurance.
It is no surprise that Cordyceps has been found to improve exercise tolerance. This is of course, how this organism has been used for centuries. The Nepalese people observed live-stock consuming Cordyceps and saw how it increased their sexual vitality and general stamina. They began to consume them as the other animals were, and found that it increased their own vitality as well – increasing stamina, endurance, and treating impotence.15
Oxidative Stress
Intense use of skeletal muscle during exercise leads to oxidative stress. Of course the human system has its own antioxidant system built in, but this system can be weakened through excessive exercise leading to more oxidative stress. Medicinal mushrooms such as Ganoderma lucidum, Tremella spp., Cordyceps spp. and Fomitopsis pinicola all help to mitigate fatigue by supporting the free radical scavenging ability within the human antioxidant system.4,6,11 These mushrooms support superoxide dismutase, glutathione reductase, and catalase; all endogenous enzymes involved in innate antioxidant functions. In a human double-blind placebo trial that assessed the oxidative stress biomarkers in athletes supplementing with Cordyceps and Ganoderma, the researchers found that after 3 months of supplementation with 1335mg Cordyceps extract per day and 1170mg Ganoderma extract per day, the athletes had significantly more free radical scavenging activity after a race than the placebo group.17
Fomitopsis pinicola
One study that explored Ganoderma tsugae, a mushroom very closely related to our local Ganoderma oregonense, found significant liver protection against exhaustive exercise-induced liver injury in rats.10 “The results concluded that G. tsugae could increase the running time to exhaustion in animals, decrease lipid peroxidation and protect against hepatic apoptosis after exhaustive exercise.” Basically, G. tsugae has protective effects on the liver that can improve exercise performance.
Blood Flow
Ganoderma spp. and Cordyceps spp. contain the nucleoside adenosine.17 Adenosine is a regulator of skeletal muscle blood flow. The role of adenosine in exercise is vascular smooth muscle relaxation, contributing to the local vasodilation which accompanies muscle contractions. Adenosine contributes around 14-29% to vasodilation in humans especially during higher frequency contractions, like during exercise. Increased 40% in exercise lasting longer than 5 minutes.2
Ganoderma applanatumalso increases endothelial nitric oxide synthase. This also has a dialing effect on the endothelial smooth muscle in our blood vessels, increasing blood flow to skeletal muscle.14
This is the first post where I have discussed mushrooms that do not necessarily grow here in the PNW. I feel confident that our local Ganoderma species would have analogous interactions with our biochemistry as Ganoderma lucidum. I also think that the west coast Hericiumspecies are analogous with Hericium erinaceus. Cordyceps are more common in the Himalayas and South America and worth investing in for personal stamina experimentation. In my experience, they are extremely powerful. We are however, extremely fortunate to have abundance of Fomitopsis pinicola, which has a beautiful relationship with our innate free-radical scavenging systems and I would propose more use for this mushroom in relation to physical exercise.
Making medicine to support you through exercise
Beet Juice Matcha Latte
Why Beets and Matcha?
Beetroot juice increases blood flow and increases efficiency of mitochondrial respiration and oxidative phosphorylation9
In a study evaluating the anti-fatigue effects of Epigallocatechin-3-gallate, a polyphenolic compound in green tea, the researchers concluded that EGCG significantly prolonged exhaustive swimming time of mice.8 In another study done on male sprinters, green tea extract supplementation prevented oxidative stress.12 The polyphenolic compounds in green tea were also found to significantly protect rats from fatigue, inflammation and tissue damage induced by acute exhaustive exercise.13
1 cup of milk of choice – I love hemp ‘milk’ straight from the teet of the hemp
½ C hot water
Directions:
Heat up milk and water
blend with all other ingredients (I use an immersion blender – you can poor into any blender, or if you have a frothing device, I imagine that would work too)
Add sweetener if necessary
Move your body!
Drink 30 min to 1 hour before working out
*If you don’t have access to powdered extracts, make a strong decoction of these mushrooms and use this as the water portion of the drink.
Geng P, Siu KC, Wang Z, Wu JY. Antifatigue Functions and Mechanisms of Edible and Medicinal Mushrooms. Biomed Res Int. 2017;2017. doi:10.1155/2017/9648496.
Ballard HJ. Invited Review ATP and adenosine in the regulation of skeletal muscle blood flow during exercise. Sheng Li Xue Bao. 2014;66(1):67-78. doi:10.13294/j.aps.2014.0009.
Mateo DC, Pazzi F, Muñoz FJD, et al. Ganoderma lucidum improves physical fitness in women with fibromyalgia . Nutr Hosp. 2015;32(5):2126-2135. doi:10.3305/nh.2015.32.5.9601.
Zhonghui Z, Xiaowei Z, Fang F. Ganoderma lucidum polysaccharides supplementation attenuates exercise-induced oxidative stress in skeletal muscle of mice. Saudi J Biol Sci. 2014;21(2):119-123. doi:10.1016/j.sjbs.2013.04.004.
Song J, Wang Y, Teng M, et al. Studies on the antifatigue activities of Cordyceps militaris fruit body extract in mouse model. Evidence-based Complement Altern Med. 2015;2015. doi:10.1155/2015/174616.
Hao L, Sheng Z, Lu J, Tao R, Jia S. Characterization and antioxidant activities of extracellular and intracellular polysaccharides from Fomitopsis pinicola. Carbohydr Polym. 2016;141:54-59. doi:10.1016/j.carbpol.2015.11.048.
Liu J, Du C, Wang Y, Yu Z. Anti-fatigue activities of polysaccharides extracted from Hericium erinaceus. Exp Ther Med. 2015;9(2):483-487. doi:10.3892/etm.2014.2139.
Teng Y, Wu D. Anti-fatigue effect of green tea polyphenols (-)-Epigallocatechin-3-Gallate (EGCG). Pharmacogn Mag. 2017;13(50):326. doi:10.4103/0973-1296.204546.
Domínguez R, Cuenca E, Maté-Muñoz JL, et al. Effects of beetroot juice supplementation on cardiorespiratory endurance in athletes. A systematic review. Nutrients. 2017;9(1):1-18. doi:10.3390/nu9010043.
Huang CC, Huang WC, Yang SC, Chan CC, Lin WT. Ganoderma tsugae hepatoprotection against exhaustive exercise-induced liver injury in rats. Molecules. 2013;18(2):1741-1754. doi:10.3390/molecules18021741.
Reis FS, Pereira E, Barros L, Sousa MJ, Martins A, Ferreira ICFR. Biomolecule profiles in inedible wild mushrooms with antioxidant value. Molecules. 2011;16(6):4328-4338. doi:10.3390/molecules16064328.
Jówko E, Długołęcka B, Makaruk B, Cieśliński I. The effect of green tea extract supplementation on exercise-induced oxidative stress parameters in male sprinters. Eur J Nutr. 2015;54(5):783-791. doi:10.1007/s00394-014-0757-1.
Liu L, Wu X, Zhang B, et al. Protective effects of tea polyphenols on exhaustive exercise-induced fatigue, inflammation and tissue damage. Food Nutr Res. 2017;61(1):1333390. doi:10.1080/16546628.2017.1333390.
Acharya K, Yonzone P, Rai M, Acharya R. Antioxidant and nitric oxide synthase activation properties of Ganoderma applanatum. Indian J Exp Biol. 2005;43(10):926-929.
Panda A, Swain K. Traditional uses and medicinal potential of Cordyceps sinensis of Sikkim. J Ayurveda Integr Med. 2011;2(1):9. doi:10.4103/0975-9476.78183.
Singh M, Tulsawani R, Koganti P, Chauhan A, Manickam M, Misra K. Cordyceps sinensis increases hypoxia tolerance by inducing heme oxygenase-1 and metallothionein via Nrf2 activation in human lung epithelial cells. Biomed Res Int. 2013;2013:1-13. doi:10.1155/2013/569206.
Rossi P, Buonocore D, Altobelli E, et al. Improving training condition assessment in endurance cyclists: Effects of ganoderma lucidum and ophiocordyceps sinensis dietary supplementation. Evidence-based Complement Altern Med. 2014;2014. doi:10.1155/2014/979613.
Fruiting bodies protrude from their hosts throughout our forests, the Ganodermas are a sight to behold and entirely hard to ignore. Ganoderma in itself means “shiny skin” of course referring to the varnished crust on many of the species in this genus, but how can we not apply this to our own, human skin. Following is research that has been done on Ganoderma lucidum, Ganoderma tsugae, and Tremella fuciformis and their uses in skin care. I am postulating that we can use out Northwest analogs, Ganoderma applanatum, Ganoderma oregonense, and Tremella mesenterica, the same way.
Sacchachitin and Polysaccharides for Wound healing
There is a product made, called Sacchachitin that is used as a wound dressing. It is made from the pulp of the Ganoderma fruiting body and when used, significantly speeds up the healing process of skin wounds. (Hung 2004) This product of course is not manufacturable by the general public, yet it is easy enough to chop up the Ganoderma into small pieces, place in a blender with a little water and create a pulp that is then simmered for about an hour. The simmering is not necessary for a styptic effect, but you want to extract the polysaccharides to see anti inflammatory, antioxidant and increased healing time effects. Speed of wound healing was also observed when Ganoderma polysaccharides were applied to the wounds of diabetic mice. It was observed that the polysaccharides accelerated the wound healing my inhibition of mitochondrial oxidative stress and improved wound angiogenesis (Tie 2012).
Healing from UVB damage
Tremella fuciformis has been used in skin care in Asia for decades, yet there is little research on our local species of Tremella, Tremella mesenterica. The polysaccharide content is comparable and so I am using the research and traditional uses of Tremella fuciformis as being analogous to the potential uses of Tremella mesenterica. Tremella is known to be a potent antioxidant and anti-inflammatory fungus. The Tremella polysaccharide extract was tested on hydrogen peroxide-induced injury of human skin fibroblasts. The polysaccharides from Tremella reduced oxidative stress and cell apoptosis in the treated skin. It also protected the skin fibroblasts from oxidative stress. (Shen, Gusman) Oxidative stress is one main reason our skin becomes wrinkled as we age, so using these polysaccharides topically could be beneficial in protecting our skin from wrinkles. The Polysaccharides, which make up about 90% of this species of mushroom, also assist the skin in its ability to retain moisture, an ability that decreases as we age. Tremella polysaccharides have also been researched for lightening skin spots in sun damaged skin and have been shown to inhibit melanin formation. Another study explored Ganoderma polysaccharides and determined that these compounds protect against “photo-aging” by eliminating UVB-induced reactive oxygen species. (Zeng 2016). One local Ganoderma to the PNW is Ganoderm oregonense, an analog to the Ganoderma Tsugae of the Eastern states. In one study, lanostane terpenoids extracted from Ganoderma tsugae fruiting bodies protected human keritinocytes from photodamage. (Lin 2013)
Triterpenoids and Polysaccharides for Atopic Dermatitis
Atopic dermatitis is a type 1 hypersensitivity reaction, which means it is an IgE mediated immediate hypersensitivity reaction, like an immediate allergic response. Researches explored a beta-glucan based cream for mild to moderate atopic dermatitis. Topical application resulted in significant improvement. In this study, the people with dermatitis put the cream on half their body, and nothing on the other half. The half of their body that the cream was applied to showed significant decline in dermatitis. (Jesenak 2015) This benefit would come from the water soluble constituents of the mushrooms, while another study looked at the lipophilic triterpenes for type 1 hypersensitivity reactions. They found that the triterpene extract inhibited histamine release from rat mast cells induced by IgE. (Rios 2010) This is a great example where a cream made from both the water and oil extract of the mushroom could be extremely beneficial for these skin conditions. Another example of a type 1 hypersensitivity reaction is the inflammation and itch that we get in response to mosquito bites. A study looked at the methanol extract of Ganoderma lucidum and the response of mosquito bitten mice. Results proved the mushroom extract to calm the scratching response of the mice. (Andoh 2010).
So Let’s Make a Body Butter and Lather ourselves With Mushroom Medicine!
This body butter is a host and fungi preparation. What this means is that the materials used are derived from both the host tree, Pseudotsuga menziesii (Doug Fir), and the mushrooms found inhabiting this tree, Ganoderma applanatum (Artist Conk) and Tremella mesenterica (Witch’s butter). The more I delve into mushroom medicine, the more I find it is important to use the tree and mushroom together in formula. So much of the energetic properties and physical properties of the mushrooms are determined by their host tree. These trees have provided oxygen, habitat and strength throughout their lifetimes in the forest and this wisdom flows through the mycelial like veins and into the mushroom fruiting body, that has now come to assist this edifice of the forest in breaking down and returning to the soil.
Materials:
1/2 C Ganoderma infused oil*
Doug Fir Pitch oil**
1Tbs cacao butter
1Tbs Shea butter
1/3oz beeswax
1tsp lanolin
1/4 C Ganoderma applanatum hydrosol or hot water extract
1/4 C Pseudotsuga menziesii hydrosol or hot water extract
1 small piece of fresh or rehydrated Tremella mesenterica
10-20 drops Doug Fir essential oil
Instruction:
Using the double boiler method, add Shea butter, Cacao butter, Ganoderma oil, Doug Fir pitch oil, and beeswax to the top bowl an melt together, stirring every so often.
In a separate jar mix the hydrosols or water extracts (make sure the extracts are at room temp if recently made. It is a good idea to make them ahead of time and keep refrigerated)
Add the Tremella piece to the mixed water solution and blend with an immersion blender until well combined.
When all the oils have melted together with the beeswax, add the lanolin (optional) and slowly poor into the jar full of the aqueous material and blend with immersion blender. After well blended, add the essential oils and blend some more. The final product should be very creamy and will become thicker after it cools off.
*Ganoderma infused oil is made my chopping up any Ganoderma species into the finest pieces possible and covering with oil, I used jojoba oil, but you can use olive oil. This is then let to sit for a few months, or I have been placing it in my dehydrator at 115 degrees F for about a week, the heat will speed up the extraction process.
**Doug Fir pitch oil is made by collecting pitch (sap/resin) from the trees and placing in a sacrificial crockpot, and covering with a small amount of oil until it is just barely covered. Let this warm for many days, strain out and you are left with a beautiful thick resinous oil.
Works Cited
Andoh, Tsugunobu, Qun Zhang, Takumi Yamamoto, Manabu Tayama, Masao Hattori, Ken Tanaka, and Yasushi Kuraishi. “Inhibitory Effects of the Methanol Extract of Ganoderma Lucidum on Mosquito Allergy Induced Itch-Associated Responses in Mice.” Journal of Pharmacological Sciences 114.3 (2010): 292-97. Web.
Gusman, Jessica Krisanti, Chien-Yih Lin, and Yang-Chia Shih. “The Optimum Submerged Culture Condition of the Culinary-Medicinal White Jelly Mushroom (Tremellomycetes) and Its Antioxidant Properties.” International Journal of Medicinal Mushrooms 16.3 (2014): 293-302. Web.
Hung, Wei-Sheng, et al. “Effect of SACCHACHITIN on Keratinocyte Proliferation and the Expressions of Type I Collagen and Tissue-Transglutaminase during Skin Wound Healing.” Journal of Biomedical Materials Research, vol. 70B, no. 1, 2004, pp. 122–129., doi:10.1002/jbm.b.30028.
Hyde, K. D., A. H. Bahkali, and M. A. Moslem. “Fungi: an Unusual Source for Cosmetics.” Fungal Diversity 43.1 (2010): 1-9. Web.
Jesenak, Milos, Slavomir Urbancek, Juraj Majtan, Peter Banovcin, and Jana Hercogova. “β-Glucan-based Cream (containing Pleuran Isolated Frompleurotus Ostreatus) in Supportive Treatment of Mild-to-moderate Atopic Dermatitis.” Journal of Dermatological Treatment 27.4 (2015): 351-54. Web.
Kurtipek, Gulcan Saylam, Arzu Ataseven, Ercan Kurtipek, Ilknur Kucukosmanoglu, and Mustafa Rasid Toksoz. “Resolution of Cutaneous Sarcoidosis Following Topical Application of Ganoderma Lucidum (Reishi Mushroom).” Dermatology and Therapy 6.1 (2016): 105-09. Web.
Rios JL. “Effects of triterpenes on the immune system”. J Ethnopharmacol. 2010;128(1):1-14
Shen, Tao, Chao Duan, Beidong Chen, Meng Li, Yang Ruan, Danni Xu, Doudou Shi, Dan Yu, Jian Li, and Changtao Wang. “Tremella fuciformis Polysaccharide Suppresses Hydrogen Peroxide-triggered Injury of Human Skin Fibroblasts via Upregulation of SIRT1.” Molecular Medicine Reports (2017): n. pag. Web.
Tie, Lu, Hong-Qin Yang, Yu An, Shao-Qiang Liu, Jing Han, Yan Xu, Min Hu, Wei-Dong Li, Alex F. Chen, Zhi-Bin Lin, and Xue-Jun Li. “Ganoderma Lucidum Polysaccharide Accelerates Refractory Wound Healing by Inhibition of Mitochondrial Oxidative Stress in Type 1 Diabetes.” Cellular Physiology and Biochemistry 29.3-4 (2012): 583-94. Web.
Zeng, Qinghai, Fang Zhou, Li Lei, Jing Chen, Jianyun Lu, Jianda Zhou, Ke Cao, Lihua Gao, Fang Xia, Shu Ding, Lihua Huang, Hong Xiang, Jingjing Wang, Yangfan Xiao, Rong Xiao, and Jinhua Huang. “Ganoderma Lucidum Polysaccharides Protect Fibroblasts against UVB-induced Photoaging.” Molecular Medicine Reports 15.1 (2016): 111-16. Web.
Some mushrooms are seen, and it is decided that there is something exclusively unique about them based on their beauty. The striking qualities of the Red Reishi with it’s varnished surface, and the Turkey Tail with its adornment of various alternating colors, have infused passerby’s with an idea that they must behold strong medicine. However, what happens when we go through life only focusing on the seemingly beautiful things, on the brightly colored splendiferous things, and do not take a moment to see the dark and explore the medicine in the seemingly unknown. There is strong medicine in the obscure, in the mushrooms, people and plants that may not show themselves luminous right away. It takes someone who is curious and who is willing to take the time to explore something deeper than surface, and to know that there is magic in everything, there is medicine everywhere, you just need to be inquisitive and unafraid of the unknown. In alchemy, preparations are made in order to extract and isolate the essentials of the organism being worked with, to uncover the ‘mistakes of nature’ and get to the core of the organism This practice teaches that there is more to all organisms than what the eye allows you to see. All things, all beings, are intended to be fully seen, and fully explored. What is seen on the surface does not usually express the crypts of our soul, and instead of looking away, we must look deeper. Same is true for seeing all living organisms, and in this case, Fomitopsis cajanderi is the chosen entity to be explored.
Distribution
Saprophytic on the dead wood of conifers and sometimes parasitic on living trees, grows most usually with others. Widely found throughout North American conifer forests.
Active known constituents
(none known, but this is what I theorize – HPLC analysis will be done on various extracts in the near future, I will amend info when that happens)
Triterpenes
Ergosterol
Beta-glucans
Phenolics
Spore print – off white
KOH – black
Therapeutic actions
cytotoxic to lymphocytic leukemia cells (in vitro), immune-modulating (most-likely), digestive bitter
Energetics
warming, sweet, tonic
Recent Research
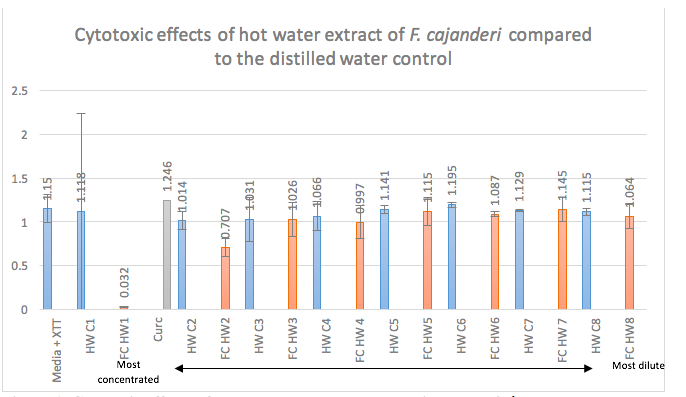
Cytotoxic Effects of Hot Ethanol Extract and Hot Aqueous Extract of Fomitopsis cajanderi on Jurkat Cell Line
Anna Sitkoff, Olivia Froehlich
INTRODUCTION
Acute lymphocytic leukemia has an incidence of about 3.4 cases per 100,000 in the United States, and each year 2,500 to 3,500 new cases of ALL are diagnosed in children. Of these cases, 15% are precursor T Lymphoblastic leukemia. Those with T cell ALL have been shown to have a high rate of remission failure and a poor overall survival as compared to B cell ALL.2,3 The Jurkat cells are a line of T lymphocytic leukemia cells. Of the new ALL cases diagnosed in children, 70-80% of them participate in clinical research trials, indicating the importance of continuing research on this cell line and potential new therapies.5
Polysaccharides and secondary metabolites produced by plants and mushrooms have been found to be a critical role in research conducted on the medicinal value of these organisms. Mushrooms are known to produce triterpene secondary metabolites and polysaccharides as an essential piece of their chitin cell wall structure. Polysaccharides and triterpenes have different, yet well researched actions on the immune system, both in vitro and in vivo. Mushroom polysaccharides, specifically beta-glucans and protein polysaccharide complexes, stimulate production of cytokines IL-12, IFN –y, and IL-2.8 IFN-y and IL-2 are of specific importance in cancer research because they stimulate natural killer (NK) cells and cytotoxic T lymphocytes (CTLs) which have antitumor effects. While polysaccharides stimulate an immune response, triterpenes directly induce cancer cell apoptosis.6 There is not yet research on Fomitopsis cajanderi, a member of the Fomitopsidaceae family, though there is some research on two other mushrooms of the same genus, Fomitopsis pinicola and Fomitopsis nigra. Fomitopside K, a lanostane triterpene glycoside extracted from F. nigra, induced apoptosis via G0/GI phase arrest in oral squamous cell carcinoma.1F. pinicola has been reported to have anti-inflammatory, antioxidant, and antimicrobial effects. F. pinicola ethanol extract has been shown to have an anticancer effect on S180 cells in vitro and in vivo, to induce ROS-dependent apoptosis, and to cause P53 mediated G1 phase arrest in human colorectal cancer cells.7 F. cajanderi is not currently vulnerable to overharvesting. The more mushrooms we know to have similar effects, the less overharvesting we do of individual species. The aim of this study is to see if this, as of yet, un-researched species of Fomitopsis, F. cajanderi, has similar medicinal properties as the other, more researched, mushrooms of this genus. The hypothesis of this particular study is that both the hot water and hot ethanol extracts of the fruiting body of F. cajanderi will show dose dependent cytotoxicity on Jurkat cells, with the hot ethanol extract showing greater cytotoxic effects.
METHODS
Collection and preparation of Fomitopsis cajanderiThe fresh Fomitopsis cajanderi was collected in January 2017 from St. Edward’s State Park in Kenmore, Washington (latitude ~47.7328° N, longitude ~122.2572° W). The mushrooms were identified and harvested by Anna Sitkoff. The mushrooms were then cleaned, chopped, dehydrated, and powdered in Bastyr University’s Botanical Medicine Lab. The mushrooms were chopped to 1cm2 pieces and weighed 195g. The specimans were placed in an Excaliber dehydrator at 46.1 degrees Celsius for 24 hours. The mushrooms were taken out of the dehydrator and weighed 64g. They were then placed in a Taiwanese grinder until a semi-powdered fibrous material was achieved. The material was stored in two separate Ziploc bags and stored in the Tierney Lab.
Preparation of hot ethanol extraction (FcETOH) The hot ethanol extraction was done in a fume hood using a reflux apparatus with a cold thumb condenser. 4g of mushroom material was weighed and placed in 250mL boiling flask with 200mL of 95% ethanol. The heating mantle was on the 5 setting to boil and then reduced to 4 with ethanol simmering for 2 hours. Refrigeration for cold water bath used for condenser was set to 12 degrees Celsius. The final extract was poured into a 50mL conical and centrifuged at 3890 RPM for 15 minutes. A .2micron steriflip filter was used to filter the extract.
Water Decoction
Dried F. cajanderi
Ethanol reflux
F. cajanderi
To concentrate the extract a rotary evaporator machine was used. The water bath was filled half way and set to 80 degrees Celsius. 90mL of FcETOH was placed into a 250mL round bottom flask, and spun hovering over the hot water bath. After 15 minutes, the flask as lowered into the bath and spun for another 15 minutes. The remaining amount of extract measured 9mL. The resulting FcETOH was 9.5% ethanol and a 1:2.25 extract. Extract was stored at 4 degrees Celsius.
Preparation of hot water decoction (FcHWE) The hot water decoction was made in Bastyr University’s Botanical Medicine Lab. The 10g F. cajanderi material was placed in 350mL of distilled water in a sterilized metal sauce pan. The decoction ran for 2 hours partially covered, 50mL more H2O was added at the beginning of the second hour. The decoction was strained using a sterilized potato-ricer and the marc was composted. The extract was then placed back on heat to reduce to 50mL for a 1:5 extract. The extract was filtered through a .2micron steriflip filter and stored at 4 degrees Celsius.
Cell culture RPMI-1640 + 10% FBS + 1:100 L-Glu media was made on 01/04/17. Jurkat cells were labeled E6.1, passage 5, cells in one vial: 5×106 cells/mL and found in Cryotank 2014, quadrant 2, cane 9, box 50, row 6, column A. They were frozen by M. Sasagawa on 10/02/2001. They were removed from the cryotank for culture on 01/23/17. Cells continued in culture to maintain cells in log growth phase. The concentration of the cells that were used for the XTT assay were 4×105 cells/mL. .1mL of cells were placed into treatment wells on a 96 U-bottom well plate.
XTT assay protocol On Day 1, a 4×105 cells/mL concentration was brought up in clear media. 100 micoliters of the cell solution was pipetted into 60 wells. 8 serial dilutions in clear media starting at a 20% concentration were made for both the FcETOH and FcHWE as well as their respective controls. The control for the FcETOH was 9.5% ETOH and the control for FcHWE was distilled H20. 0.1mL of extract and control dilutions were added to the 0.1mL cells in triplicates. 0.1mL of 2% solution of 50mmol curcumin/dmp was also added as a positive cytotoxic control in triplicates. 200 microliters of clear media were placed in each of the surrounding wells to act as a evaporation buffer. Plate was placed in the incubator set at 37 degrees Celsius and 5% CO2.
Day 2—continued incubation.
On Day 3, the plate was spun at 750 rpm for 5 minutes. The media was aspirated off of the 6×11 block of wells using a micropipette tip on glass tube vacuum. Cells were rinsed by adding 200 microliters of PBS to the 6×11 block of wells and spun at 750 rpm for 5 minutes. PBS was aspirated off and the rinse was repeated once more for a total of two cell rinsings. The 200 microliters of clear media were aspirated from each of the surrounding wells and replaced with 100 microliters of fresh clear media. The XTT assay solution was made and 100 microliters of it were added to each the wells within the 6×11 block. The plate incubated for 3 hours. After this time, the plate was covered with foil to avoid light reaction and placed on an agitator for 15 minutes. It was then read on a microplate reader using SoftMax Pro.
RESULTS
XTT assay interpretation is based on light absorbency with lower values indicating less live cells present (i.e. greater cytotoxic effects). Figure 1 shows increasing cytotoxicity from FcHWE2, with FcHWE1 having the greatest cytotoxicity when compared to the control. Figure 2 shows dose dependent cytotoxicity of the FcETOH when compared to the control. See conclusion for explanation on absorbency readings of FcETOH1-4.
DISCUSSION
Positive cell death control of curcumin Reading of an XTT assay involves measuring light or color absorbency at 492 nm and 650 nm with higher values, or more color present, indicating viable cells. Curcumin is bright yellow, which poses a problem for a cell death/cell viability assay that relies on light or color absorbency for its values. The cells were rinsed with PBS and spun twice in an attempt to remove as much of the curcumin as possible and to avoid false readings. However, Figures 1 and 2 both demonstrate that curcumin had the highest light or color absorbency, which is exactly opposite of what is expected for positive cell death. It is suspected that there was some sort of cell death by the curcumin, but its residual color produced high values during the analysis of the XTT assay. This being said, there is essentially a lack of a genuine reading for known positive cell death for this experiment. In future research involving an XTT assay, a different, colorless positive cell death control should be used.
Absorbency of the FcETOH preparation at increasing concentration Similar to the issue regarding the positive cell death control of curcumin, the FcETOH extract initially showed lower XTT values starting at the 2nd most dilute preparation (cytotoxicity), but then showed gradually higher XTT values started at FcETOH5 (Figure 2). While preparing the plates, it was noted that the three highest concentration preparations of FcETOH reacted with the clear media containing the Jurkat cells upon combination, resulting in a cloudy solution. This was observed in the first XTT assay (not discussed). In an attempt to prevent this cloudiness from interfering with the light absorbency of the XTT assay, the cells were rinsed with PBS and spun twice. Figure 2 shows that there was gradual increasing absorbency starting at FcETOH4, a finding that supports the thought that the cloudiness would interfere with the analysis. During the two cell rinses, the presence of cell pellets were not visible for the three highest FcETOH concentrations as they were for the blanks and the ETOH control wells. Therefore, it is likely possible that the highest concentrations of FcETOH had continually or completely effective cytotoxicity on the Jurkat cells, but the reaction between the preparation and media prevented the XTT assay from supporting these claims.
CONCLUSION
There were dose dependent cytotoxic effects of both the FcETOH and FcHWE preparations compared to their respective controls. FcETOH showed higher dose dependent cytotoxicity than FcHWE preparations. This result is concurrent with the hypothesis that the hot ethanol extract would have more direct cytotoxicity because of the triterpene extraction (triterpenes have been found to have a direct cytotoxic effect6,7) typical of most hot ethanol mushroom extracts. Further research needs to be done exploring the direct mechanism of cytotoxicity with this specific mushroom extract. Most of what is known of aqueous extracts involves their extraction of polysaccharides which are typically indirectly cytotoxic by inducing cytokine production. Therefore, further studies need to explore if the FcHWE cytotoxic effect is direct or from stimulating IFN-y and IL-2 production. HPLC analysis should be done on both the FcETOH and FcHWE to explore specific constituents of F. cajanderi and specifically to determine if it contains the cytotoxic lanostane triterpene glycosides as related species do.
Appendix
Figure 1. Cytotoxic effects of FcHWE on Jurket cells starting at the 2nd most concentrated preparation. Jurkat cells were treated with triplicates of 8 serial dilutions of FcHWE and its distilled water control for 48 hours. XTT assay solution was added and incubated for 3 hours. Absorbency was read at 650 nm and 492 nm with higher values indicating cell viability. Numbers displayed correspond to respective absorbency values.
Figure 2. Cytotoxic effects of FcETOH on Jurket cells starting at the 2nd most dilute preparation. Jurkat cells were treated with triplicates of 8 serial dilutions of FcETOH and its 9.5% ETOH control for 48 hours. XTT assay solution was added and incubated for 3 hours. Absorbency was read at 650 nm and 492 nm with higher values indicating cell viability. Numbers displayed correspond to respective absorbency values.
Figure 3. Comparison of cytotoxic effects of FcETOH and FcHWE with FcETOH being having increased cytotoxic effects. Both plates for FcETOH and FcHWE were prepared and read in the same fashion. The FcETOH showed dramatically decreased absorbency starting at the 2nd most dilute preparation while the FcHWE only started showing decreased absorbency at the 2nd most concentrated preparation. Numbers displayed correspond to respective absorbency values.
Work Cited
Bhattarai G, Lee Y-H, Lee N-H, et al. Fomitoside-K from Fomitopsis nigra Induces Apoptosis of Human Oral Squamous Cell Carcinomas (YD-10B) via Mitochondrial Signaling Pathway. Biol Pharm Bull. 2012;35(10):1711-1719. doi:10.1248/bpb.12-00297.
Coustan-Smith E, Mullighan CG, Onciu M, et al. Early T-cell precursor leukaemia: a subtype of very high-risk acute lymphoblastic leukaemia. Lancet Oncol. 2009;10(2):147-156. doi:10.1016/S1470-2045(08)70314-0.
Inukai T, Kiyokawa N, Campana D, et al. Clinical significance of early T-cell precursor acute lymphoblastic leukaemia: Results of the Tokyo Children’s Cancer Study Group Study L99-15. Br J Haematol. 2012;156(3):358-365. doi:10.1111/j.1365-2141.2011.08955.x.
Lee IK, Jung JY, Yeom JH, et al. Fomitoside K, a new lanostane triterpene glycoside from the fruiting body of Fomitopsis nigra. Mycobiology. 2012;40(1):76-78. doi:10.5941/MYCO.2012.40.1.076.
Pui C-H, Yang JJ, Hunger SP, et al. Childhood Acute Lymphoblastic Leukemia: Progress Through Collaboration. J Clin Oncol. 2015;33(27):JCO.2014.59.1636. doi:10.1200/JCO.2014.59.1636.
Ríos JL. Effects of triterpenes on the immune system. J Ethnopharmacol. 2010;128(1):1-14. doi:10.1016/j.jep.2009.12.045.
Wang Y, Cheng X, Wang P, et al. Investigating migration inhibition and apoptotic effects of Fomitopsis pinicola chloroform extract on human colorectal cancer SW-480 cells. PLoS One. 2014;9(7):1-13. doi:10.1371/journal.pone.0101303.
Wasser S. Medicinal mushrooms as a source of antitumor and immunomodulating polysaccharides. Appl Microbiol Biotechnol. 2003;60(3):258-274. doi:10.1007/s00253-002-1076-7.