Ergosterol is as ubiquitous in mushrooms as cholesterol is in humans. It is formed by an almost identical metabolic process, the mevalonate pathway. When mushrooms are exposed to ultraviolet light, ergosterol is converted to ergocalciferol, or vitamin D2. Once ingested, vitamin D2 is converted to calcidiol in the liver and eventually calcitriol, our active vitamin D3, in the kidneys. There is clinical evidence demonstrating that ergocalciferol-D2, although not as bioavailable as calcitriol, could improve vitamin D deficiency in humans (1,2,3).
Mushrooms are an excellent source of vitamin D2, particularly if they have been exposed to ample UV light (4). Researchers found that when people consumed chanterelle mushrooms containing 14 mcg of ergocalciferol with their lunch, that their serum vitamin D levels improved as much as the group who was receiving ergocalciferol supplementation (5). However, another study found that vitamin D levels decreased significantly after cooking mushrooms and serum concentrations of vitamin D after consumption of 100 g fresh sliced cooked mushrooms for 16 weeks did not show significant improvement in serum vitamin D levels. This was, notably, in a prediabetic, overweight population (6). In healthy young adults, serum vitamin D levels did improve after consumption of vitamin D2-enhanced button mushrooms via UV-B irradiation and to a similar degree as with a vitamin D2 supplement (7).
The benefits of optimal serum vitamin D levels include and are not limited to:
Bone health, kidney health, immune modulation, calcium homeostasis, and cardiovascular health (8).
Vaes AMM, Tieland M, de Regt MF, Wittwer J, van Loon LJC, de Groot LCPGM. Dose-response effects of supplementation with calcifediol on serum 25-hydroxyvitamin D status and its metabolites: A randomized controlled trial in older adults. Clin Nutr. 2018;37(3):808-814. doi:10.1016/j.clnu.2017.03.029
Martineau AR, Thummel KE, Wang Z, et al. Differential Effects of Oral Boluses of Vitamin D2 vs Vitamin D3 on Vitamin D Metabolism: A Randomized Controlled Trial. J Clin Endocrinol Metab. 2019;104(12):5831-5839. doi:10.1210/jc.2019-00207
Seijo M, Mastaglia S, Brito G, Somoza J, Oliveri B. Es equivalente la suplementación diaria con vitamina D2 o vitamina D3 en adultos mayores? [Is daily supplementation with vitamin D2 equivalent to daily supplementation with vitamin D3 in the elderly?]. Medicina (B Aires). 2012;72(3):195-200.
Kamweru PK, Tindibale EL. Vitamin D and Vitamin D from Ultraviolet-Irradiated Mushrooms (Review). Int J Med Mushrooms. 2016;18(3):205-214. doi:10.1615/IntJMedMushrooms.v18.i3.30
Outila TA, Mattila PH, Piironen VI, Lamberg-Allardt CJ. Bioavailability of vitamin D from wild edible mushrooms (Cantharellus tubaeformis) as measured with a human bioassay. Am J Clin Nutr. 1999;69(1):95-98. doi:10.1093/ajcn/69.1.95
Mehrotra A, Calvo MS, Beelman RB, et al. Bioavailability of vitamin D2 from enriched mushrooms in prediabetic adults: a randomized controlled trial. Eur J Clin Nutr. 2014;68(10):1154-1160. doi:10.1038/ejcn.2014.157
Urbain P, Singler F, Ihorst G, Biesalski HK, Bertz H. Bioavailability of vitamin D₂ from UV-B-irradiated button mushrooms in healthy adults deficient in serum 25-hydroxyvitamin D: a randomized controlled trial. Eur J Clin Nutr. 2011;65(8):965-971. doi:10.1038/ejcn.2011.53
Charoenngam N, Holick MF. Immunologic Effects of Vitamin D on Human Health and Disease. Nutrients. 2020;12(7):2097. Published 2020 Jul 15. doi:10.3390/nu12072097
Terpenes are fat- and alcohol-soluble compounds that come in many variations – monoterpenes, sesquiterpenes, diterpenes and triterpenes. The prefix indicates the number of carbons in the molecule: 10, 15, 20, and 30, respectively. As the terpene becomes bigger, it also becomes heavier and less volatile. The mushroom smell many are familiar with is in part due to low-molecular weight molecules that make their way into the nose – the monoterpenes and sesquiterpenes. These lower-weight chemicals act as attractants and deterrents to insects and other invertebrates, acting as both pheromones and defense chemicals. The most well-known terpenes in fungi are both found in lion’s mane. Erinicines, a group of diterpenoids, have nootropic properties and are found in lion’s mane mycelium, while the terpene hericenones, which are also nootropic, are found in lion’s mane fruiting body.
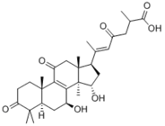
As the mushroom matures from primordia to fruiting body, an enzyme stimulates the synthesis of triterpenes. Because of this, there are significantly more triterpenes in the mushroom compared to the mycelium and primordia. Triterpenes in mushrooms are lanostane triterpenes, and can be referred to as “lanostanoids”. The most well-known triterpenes found in mushrooms are ganoderic acids, found in various species of reishi. These terpenes are major compounds of interest in cancer research because they have cytotoxic activity in many cancer cell lines; they also demonstrate specific antiviral activity, and are helpful in the treatment of hypersensitivity reactions.
Ganoderic acid, a triterpene isolated from Ganoderma spp.
Anti-neoplastic
There are a significant number of in vitro and in vivo animal studies demonstrating direct anti-neoplastic activity of triterpenes. These molecules are cytotoxic to many cancer cell lines, as well as instigators of an antineoplastic immune response (1,2,3,4,5,6).
Anti-viral
Mushroom-derived triterpenes and sesquiterpenes have specific antiviral activity through the inhibition of the enzyme neuraminidase, which viruses use to leave one cell to enter another (7,8). In addition to anti-neuraminidase activity, triterpenes stimulate an immune response that preferences cytotoxic CD8 T cells, a major component of adaptive antiviral immunity (9).
Anti-allergic
Triterpenes may be very beneficial in the treatment of hypersensitivity reactions including allergy, dermatitis and rhinitis. Ganoderic acids from reishi inhibit histamine release from mast cells, which can moderate the severity of allergic reactions (10). Ganoderic acids have also been shown to induce the release of the cytokines IL-10 and IFN-𝛾, suppressing TH2 responses in favor of TH1, and moving the immune system away from allergic responses (11).
Neuroprotective
Erinacines and hericenones, diterpenes found in lion’s mane mycelium and fruiting body, respectively, both have neuroprotective activity (12). Both molecules have demonstrated neurogenesis via nerve growth factor synthase activation, and erinacines improve plaque formation in mouse models of Alzheimer’s disease while increasing hippocampal neurogenesis (12).
Absorption
Only 10% of terpenes found in fungi are bioavailable with oral consumption, potentially due to first-pass metabolism in the liver and metabolism by intestinal bacteria (13). Raw mushrooms offer particularly poor bioavailability, as terpenes remain trapped inside fungal cells, while mushroom extracts offer improved absorption. Taking terpene-rich extracts with food impairs absorption; for best results, take terpene-rich mushrooms as extracts on an empty stomach (14). To learn more, check out my blog post on factors impacting the absorption of mushroom constituents.
DIY Extraction of Terpenes
Terpenes are alcohol- and fat-soluble and range from low- to high-molecular weight
Infuse chopped mushrooms into oil and use as a base for lotion, salad dressings or flavorful drizzle
Soak dried and chopped mushrooms in alcohol as part of a double extraction process.
1. Ríos JL. Effects of triterpenes on the immune system. J Ethnopharmacol. 2010;128(1):1-14. doi:10.1016/j.jep.2009.12.045.
2. Grienke U, Kaserer T, Pfluger F, et al. Accessing biological actions of Ganoderma secondary metabolites by in silico profiling. Phytochemistry. 2015;114:114-124. doi:10.1016/j.phytochem.2014.10.010.
3. Bhattarai G, Lee Y-H, Lee N-H, et al. Fomitoside-K from Fomitopsis nigra Induces Apoptosis of Human Oral Squamous Cell Carcinomas (YD-10B) via Mitochondrial Signaling Pathway. Biol Pharm Bull. 2012;35(10):1711-1719. doi:10.1248/bpb.12-00297.
4. Ren G, Liu XY, Zhu HK, Yang SZ, Fu CX. Evaluation of cytotoxic activities of some medicinal polypore fungi from China. Fitoterapia. 2006;77(5):408-410. doi:10.1016/j.fitote.2006.05.004.
5. Jin X, Ruiz Beguerie J, Sze DM-Y, Chan GCF. Ganoderma lucidum (Reishi mushroom) for cancer treatment. Cochrane database Syst Rev. 2016;4:CD007731. doi:10.1002/14651858.CD007731.pub3.
6. Zhu Q, Bang TH, Ohnuki K, Sawai T, Sawai K, Shimizu K. Inhibition of neuraminidase by Ganoderma triterpenoids and implications for neuraminidase inhibitor design. Sci Rep. 2015;5(AUGUST):13194. doi:10.1038/srep13194.
7. Teplyakova T V., Psurtseva N V., Kosogova TA, Mazurkova NA, Khanin VA, Vlasenko VA. Antiviral Activity of Polyporoid Mushrooms (Higher Basidiomycetes) from Altai Mountains (Russia). Int J Med Mushrooms. 2012;14(1):37-45. doi:10.1615/IntJMedMushr.v14.i1.40.
8. Gao L, Sun Y, Si J, et al. Cryptoporus volvatus extract inhibits influenza virus replication in vitro and in vivo. PLoS One. 2014;9(12). doi:10.1371/journal.pone.0113604.
9. Van Thu Nguyen, Nguyen The Tung, To Dao Cuong, Tran Manh Hung, Jeong Ah Kim, Mi Hee Woo, Jae Sue Choi, Jeong-Hyung Lee, Byung Sun Min, Cytotoxic and anti-angiogenic effects of lanostane triterpenoids from Ganoderma lucidum, Phytochemistry Letters, Volume 12, 2015, Pages 69-74, ISSN 1874-3900,
10. Bhardwaj N, Katyal P, Sharma AK. Suppression of inflammatory and allergic responses by pharmacologically potent fungus Ganoderma lucidum. Recent Pat Inflamm Allergy Drug Discov. 2014;8(2):104-17. doi: 10.2174/1872213×08666140619110657. PMID: 24948193.
11. Liu C, Cao M, Yang N, Reid-Adam J, Tversky J, Zhan J, Li XM. Time-dependent dual beneficial modulation of interferon-γ, interleukin 5, and Treg cytokines in asthma patient peripheral blood mononuclear cells by ganoderic acid B. Phytother Res. 2022 Mar;36(3):1231-1240. doi: 10.1002/ptr.7266. Epub 2022 Feb 3. PMID: 35112740.
12. Friedman M. Chemistry, Nutrition, and Health-Promoting Properties of Hericium erinaceus (Lion’s Mane) Mushroom Fruiting Bodies and Mycelia and Their Bioactive Compounds. J Agric Food Chem. 2015;63(32):7108-7123. doi:10.1021/acs.jafc.5b02914
13. Teekachunhatean, S., Sadja, S., Ampasavate, C., Chiranthanut, N., Rojanasthien, N., & Sangdee, C. (2012). Pharmacokinetics of Ganoderic Acids A and F after Oral Administration of Ling Zhi Preparation in Healthy Male Volunteers. Evidence-Based Complementary and Alternative Medicine, 2012, 780892. https://doi.org/10.1155/2012/780892
14. Verhoeven J, Keller D, Verbruggen S, Abboud KY, Venema K. A blend of 3 mushrooms dose-dependently increases butyrate production by the gut microbiota. Benef Microbes. 2021;12(6):601-612. doi:10.3920/BM2021.0015
Polysaccharides are a major component of the fungal cell wall and possibly the most potent and abundant therapeutic constituent in mushroom medicine. They are well-touted for their immune regulatory activity, but they play a role in many other important physiologic processes including blood sugar regulation, inflammation modulation, antioxidant activity, mood support, and modulation of the intestinal microbiome and the greater intestinal environment.
Biologic Response Modifiers
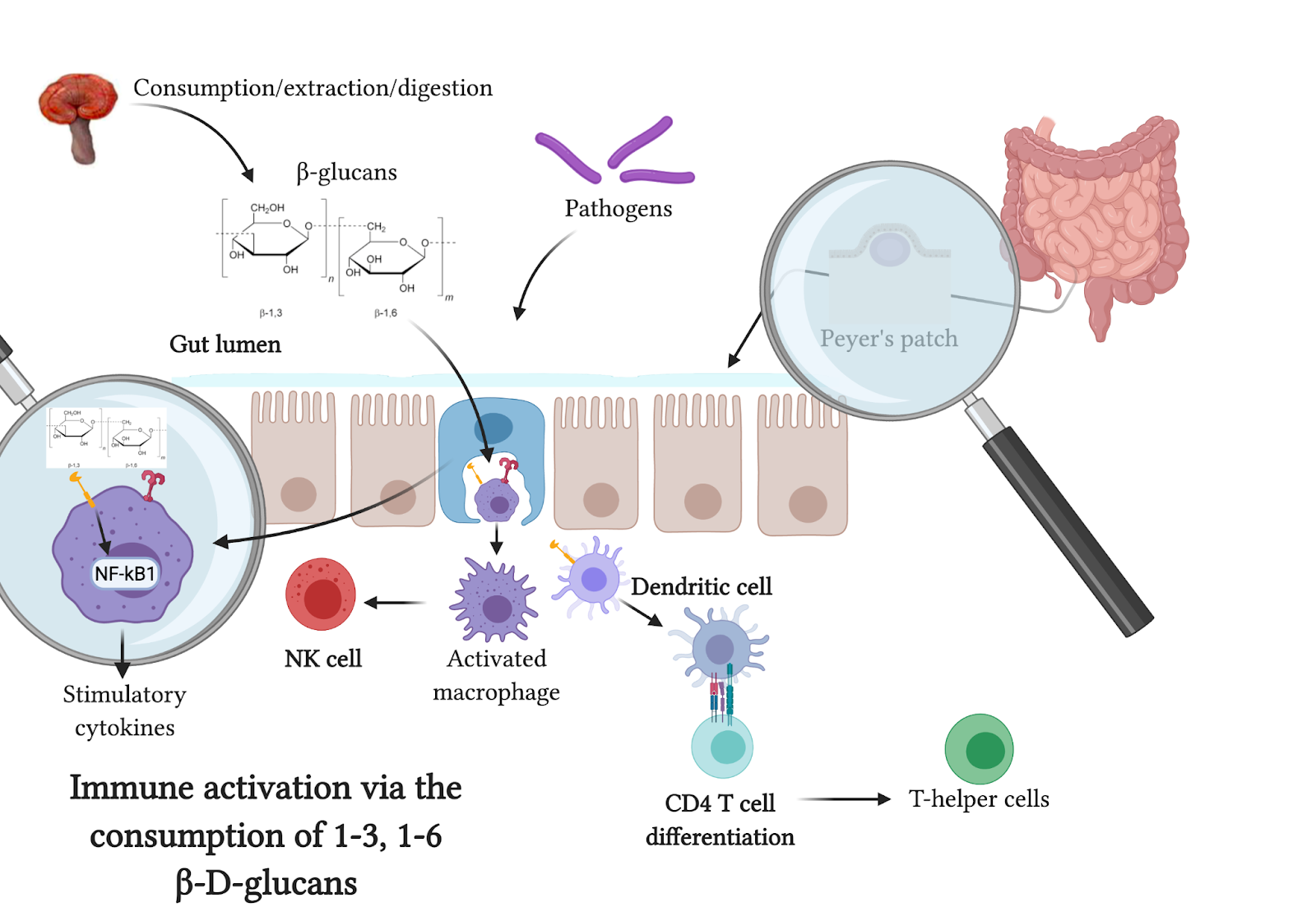
The fungal cell wall is composed of many layers – mannoproteins, ß- (1,6)-glucan and ß-(1,3)-glucan, and chitin. Humans do not make chitinase, and cannot utilize chitin for anything other than non-digestible fiber. Chitin can, however, be broken down by different processing techniques, including heat, allowing the mannan and beta-glucan layers to be utilized as medicine. From an immunological perspective, these layers are considered pathogen-associated molecular patterns (PAMPs), and bind to pathogen recognition receptors (PRRs) on cells in the innate immune system – macrophages, granulocytes, dendritic cells and natural killer cells. The majority of binding occurs in the gut-associated lymphoid tissue (GALT), where Peyer’s patches contain cells of the innate immune system. Specific receptors that bind these ß-glucans include Dectin-1 and TLR-2 (1). Binding has a hormetic effect, creating a stress response in our immune system, stimulating the release of cytokines with anti-inflammatory, pro-inflammatory, and other immune-modulating actions. These cytokines stimulate an increase in natural killer cells and cytotoxic T cells, generating a stress signal that leads to a more responsive immune system.
Blood Sugar regulation
ß-glucan polysaccharides from mushrooms inhibit the enzyme alpha-glucosidase, which is essential for carbohydrate metabolism and for generating a rise in blood sugar following a meal. They have also been shown to decrease gastric emptying, slowing the absorption of glucose, and preventing hyperglycemic episodes. Finally, polysaccharides demonstrate the ability to upregulate GLUT 4, an insulin-response glucose transporter, while down-regulating NF-κB, a nuclear transcription factor that regulates inflammation (2,3).
Antioxidant
ß-glucan polysaccharides increase the activity of hepatic oxidative enzymes such as catalase, glutathione peroxidase, and superoxide dismutase, increasing levels of glutathione and decreasing malondialdehyde levels (a marker of lipid peroxidation). Generally, these compounds support our innate antioxidant systems, relieving oxidative stress, and reducing free radical damage (4).
Prebiotic
The oligosaccharides and polysaccharides contained in mushrooms are consumed by Bifidobacterium and Lactobacilli, two important bacterial phyla in a healthy microbiome (5,6). Polysaccharides also promote production of important postbiotics like butyric acid, a short chain fatty acid, which provides colon cells with about 70% of their total energy needs (7).
Psychobiotic
Polysaccharides play a role in creating a healthy intestinal ecosystem through potentiating the growth of healthy bacteria, archaea and fungi while also supporting a more resilient gastrointestinal terrain. Healthy gut flora and intestinal mucosa are associated with better mood. Signals from the GI tract can independently relay information to the central nervous system, and the maintenance of a healthy GI tract supports a balanced central nervous system. There are a number of studies that have demonstrated increased probiotic intake associated with increased mood and diminished anxiety (8). Specifically, probiotic supplementation with Lactobacillus helveticus and Bifidobacterium longum showed less self-reported negative mood and decreased urinary cortisol. A similar effect was also observed in healthy participants who consumed a mixture of Bifidobacterium bifidum and Bifidobacterium lactis, and Lactobacillus acidophilus, Brevibacillus brevis, Brevibacterium casei, Bifidobacterium salivarius, and Lactococcus lactis (9,10).
Absorption
Raw mushroom preparations (including products labeled “mushroom powder”) have not undergone sufficient processing to break down the chitin in the cell wall, leaving ß-glucans unavailable for absorption; extractions employ heat, water, and alcohol to free ß-glucans, improving their absorption and activity. While mixing mushrooms with coffee, tea, or chocolate is increasingly popular, the tannins in these foods bind the fungal polysaccharides, diminishing bioavailability (11). For best results, take polysaccharide-rich mushroom preparation on an empty stomach, away from food. To learn more, check out my blog post on factors impacting the absorption of mushroom constituents.
DIY Extraction of Polysaccharides
Polysaccharides are water soluble and high molecular weight (heavy molecules that don’t evaporate)
Boil chopped (fresh or dried) mushrooms in water for about 2 hours. Use this water as a base for creams, teas, stews, soups and concentrated extracts
References
Baert K, Sonck E, Goddeeris BM, Devriendt B, Cox E. Cell type-specific differences in β glucan recognition and signalling in porcine innate immune cells. Dev Comp Immunol. 2015;48(1):192-203. doi:10.1016/j.dci.2014.10.005.
Vitak T, Yurkiv B, Wasser S, Nevo E, Sybirna N. Effect of medicinal mushrooms on blood cells under conditions of diabetes mellitus. 2017;8(5):187-201. doi:10.4239/wjd.v8.i5.187.
Chen Y, Liu Y, Rahman M, Yan X, Yang C. Structural characterization and antidiabetic potential of a novel heteropolysaccharide from Grifola frondosa via IRS1 / PI3K-JNK signaling pathways. Carbohydr Polym. 2018;198(15):452-461. doi:10.1016/j.carbpol.2018.06.077.
Wang H, Liu YM, Qi ZM, et al. An Overview on Natural Polysaccharides with Antioxidant Properties. 2015;(February). doi:10.2174/0929867311320230006.
Yu Z, Liu B, Mukherjee P, Newburg DS. Trametes versicolor Extract Modifies Human Fecal Microbiota Composition In vitro. 2013:107-112. doi:10.1007/s11130-013-0342-4.
Palacios S, Losa F, Dexeus D, Cortés J. Beneficial effects of a Coriolus versicolor – based vaginal gel on cervical epithelization , vaginal microbiota and vaginal health : a pilot study in asymptomatic women. 2017:4-9. doi:10.1186/s12905-017-0374-2.
Verhoeven J, Keller D, Verbruggen S, Abboud KY, Venema K. A blend of 3 mushrooms dose-dependently increases butyrate production by the gut microbiota. Benef Microbes. 2021 Nov 16;12(6):601-612. doi: 10.3920/BM2021.0015. Epub 2021 Sep 30. PMID: 34590532.
Messaoudi M, Violle N, Bisson J-F, Desor D, Javelot H, Rougeot C. Beneficial psychological effects of a probiotic formulation (Lactobacillus helveticus R0052 and Bifidobacterium longum R0175) in healthy human volunteers. Gut Microbes 2011;2:256– 261.
Steenbergen L, Sellaro R, van Hemert S, Bosch JA, Colzato LS. A randomized controlled trial to test the effect of multispecies probiotics on cognitive reactivity to sad mood. Brain Behav Immun 2015;48:258–264.
Allen AP, HutchW, Borre YE, et al. Bifidobacterium longum 1714 as a translational psychobiotic: modulation of stress, electrophysiology and neurocognition in healthy volunteers. Transl Psychiatry 2016;6:e939.
Li R, Zeng Z, Fu G, Wan Y, Liu C, McClements DJ. Formation and characterization of tannic acid/beta-glucan complexes: Influence of pH, ionic strength, and temperature. Food Res Int. 2019;120:748-755. doi:10.1016/j.foodres.2018.11.034
Proving that mushrooms are an essential part of the human diet
Definitions to consider while reading:
Antioxidant: Compounds that mitigate oxidation. Oxidation is a reaction that can produce free radicals, leading to cellular damage. Antioxidants such as thiols and ascorbic acid (vitamin C) terminate these reactions by donating electrons. To balance oxidative states, living organisms maintain systems with overlapping antioxidants such as glutathione, catalase, superoxide dismutase.
Blood-brain barrier: a filtering mechanism of the capillaries that carry blood to the brain and spinal cord tissue, blocking the passage of certain substances.
Cation chelator: a type of bonding of ions and molecules to metal ions. There is application of this in removing toxic metals from the body.
Glutathione: Glutathione is a tripeptide (cysteine, glycine, and glutamic acid) found in surprisingly high levels—5 millimolar—concentrations in most cells. Responsible for the following actions in the body: (Pizzorno)
Direct chemical neutralization of singlet oxygen, hydroxyl radicals, and superoxide radicals
Cofactor for several antioxidant enzymes
Regeneration of vitamins C and E
Neutralization of free radicals produced by Phase I liver metabolism of chemical toxins
One of approximately 7 liver Phase II reactions, which conjugate the activated intermediates produced by Phase I to make them water soluble for excretion by the kidneys
Transportation of mercury out of cells and the brain
Regulation of cellular proliferation and apoptosis
Vital to mitochondrial function and maintenance of mitochondrial DNA (mtDNA)
Lipid peroxidation: The oxidative degradation of lipids. It is the process in which free radicals “steal” electrons from the lipids in cell membranes, resulting in cell damage. (Wik)
Oxidative stress: disturbance in the balance between the production of reactive oxygen species (free radicals) and antioxidant defenses (Betteridge)
Pleurotus ostreatus – Oyster Mushroom
‘Constituentism’
I am not necessarily a fan of isolation and ‘constituentism’ in supplemental medicine – especially when a constituent has been isolated from its synergistic community of molecules from the whole plant or whole mushroom and is considered supposedly different than a pharmaceutical. Though, I do love to understand the individual constituents in organisms and how they work independently, in order to better understand how they work synergistically. There is also a greater understanding to be had of how animals, plants, fungi and bacteria all evolved together when we can explore specific plant and fungi constituent mechanisms with specific cellular receptors and transporters in the animal organism. A specific example of this is a compound called Ergothioneine (ERG). ERG is an amino acid derivative, specifically a crystalline betaine, derivative of histidine – the etymology explains; ergot- (found in ergot (fungus) thione- (double bond to a Sulphur) and -ine (amine containing)—basically a sulphur containing amine found in fungi. ERG is also found in Actinobacteria (gram positive bacteria that behave similar to fungi in soil, helping to decompose the organic matter of dead organisms so the molecules can be taken up by new plants, they even grow extensive mycelium like fungi and were in fact long believed to be fungi) and Cyanobacteria, commonly known as blue-green algea, (another abnormal bacteria that can actually obtain their energy from photosynthesis – they are the only photosynthetic prokaryote able to produce oxygen). (blankenfeldt)
Specific ERG transporters in the animal
We have evolved so closely with mushrooms that we have specific transporters and receptors uniquely for mushroom compounds. ERG is no exception. While we have a specific receptor for ERG, we cannot synthesize this molecule ourselves, and as I will describe later, we are discovering this compound is extremely important for human health. The ERG transporter, OCTN1, (gene: SLC22A4) is found specifically on erythrocytes, fetal liver and bone marrow, ileum of the small intestine, trachea, kidney, cerebellum, lung, monocytes, seminal vesicles and the lens and cornea of the eye (Halliwell). Interestingly, ERG transporter is concentrated in the mitochondria of cells – suggesting a role in protecting mitochondrial components from DNA damage (Paul, Halliwell). This transporter’s only known role is to sequester as much ERG within the cells as is available, and only cells with this transporter can absorb, distribute, and retain this compound (Grundermann). Researchers explored the effects when cells were depleted of this transporter, the results being that cells were more susceptible to oxidative stress, leading to mitochondrial damage, protein oxidation, and lipid peroxidation. Once taken up into the cell, ERG is extremely bioavailable and is retained for up to a month within the body.
Bioavailability of ergothioneine from mushroom consumption
Once consumed in the diet, whether it be the isolated molecule or from whole mushrooms, ERG is quickly absorbed into the blood stream. Human erythrocytes (red blood cells, RBC) contain 2-9 fold more ERG than plasma. Interestingly, concentration is lowest in early life (1-10yo), increases between 11-18yo and reaches maximum value of 3.7mg/100mL by age 18. In this study, subjects consumed 8g and 16g of mushrooms and increases in red blood cell ERG were observed. After 1 and 4hr of consumption, the 16g mushroom dose increased RBC ERG concentration compared to the control, and after only 2 hrs of 16g of mushroom consumption, RBC ERG concentration was significantly higher than the control. Variability in bioavailablity was due to genetic variations in the SLC22A4 gene (Heller, Kris-etherton, Beelman). Interestingly, mutations in the ERG transporter have been identified as a susceptibility factor for autoimmune disorders, such as rheumatoid arthritis and Crohn’s disease, as well as neurodegenerative disorders, such as dementia and Parkinson’s (Paul, Snyder).
Why do we care – Once ergothioneine is in the cell, then what happens?
Ergothioneine as an antioxidant and cytoprotectant: The distribution of ERG transporters may seem random, but on closer inspection, they seem to be present in tissues predisposed to high levels of oxidative stress and inflammation (Halliwell). In animal studies, animals who were completely deficient in ERG had higher levels of reactive oxygen species and were therefore more susceptible to oxidative stress. In vitro, ERG is a powerful scavenger of hydroxyl radical and has been shown to deactivate singlet oxygen at a higher rate than glutathione (Hseu, Servillo). Similar results were proven in vivo – rats supplemented with ERG had lower levels of lipid peroxidation and higher levels of glutathione and alpha-tocopherol. As one might surmise, ERG and glutathione seem to have an intimate relationship within the cell. Glutathione is considered the major intracellular antioxidant in almost all organisms and has important functions in detoxification and immune function. It has been proposed, based on current research, that ERG can help maintain GSH levels in the presence of oxidative burden by interacting with other cellular defense systems. The maintenance of glutathione tissue levels is important in maintaining health as depletion will impair immune function. Conveniently, mushrooms contain both glutathione and ERG. In fact, mushrooms have been observed to have higher GSH amounts than any vegetable or fruit. Grifola frondosa (Maitake), Hericium erinaceus (Lions mane), Pleurotus ostreatus (Oyster mushroom), Boletus edulis (porcini) and Lentinus edodes (shitake) containing the most, respectively (Kalaras). Considering the relationship between ERG and GSH in mushrooms, a correlation analysis found that mushrooms high in GSH were also high in ERG, specifically the caps or pileus of the mushrooms (kalaras). All this said, mushrooms are an important source for cellular antioxidants.
Cation chelator: ERG chelates divalent metal cations – specifically, Cu2, hg, Zn, Cd, Co, Fe, and Ni (Cheah, Song, Kerley). Binding these cations in the body may help prevent their participation in the generation of reactive oxygen species. For example, EGT has been found to protect DNA and protein against copper induced oxidative damage through formation of a redox-inactive EGT-Cu complex. Interestingly, the high levels of ERG in semen – due to the high concentration of ERG OCTN1 transporter on the seminal vesicles- have been shown to prevent Cu inhibition of sperm motility. There is more to be explored between the relationship of ERG and semen viability.
Ergothioneine, aging, and cognitive decline: As we age, we are more vulnerable to the oxidative stress and environmental toxins that slowly damage our DNA, making us more susceptible to neurodegeneration – one of many conditions associated with DNA damage from oxidative stress. Low levels of glutathione have been linked to certain neurodegenerative diseases including Parkinson’s disease (Wei, Shah). ERG was able to dose dependently enhance glutathione activity in the rat liver cytosol—leading researchers to postulate that declining ERG may play a role in age related decline of GSH and glutathione peroxidase. Furthermore, ERG levels were found to be lower in the elderly with early stages of dementia and in PD patients relative to age matched healthy controls (Cheah). In animal studies, oral administration of ERG protected neurons and preserved cognitive function following administration of toxic amyloid beta cisplatin or D-galactose. It is now understood that the presence of Octn1 transporter in the blood brain barrier is responsible for these neuroprotective actions. Captivatingly, researchers found that there is a direct relationship between whole blood and brain ERG levels following consumption (Cheah). It is rare for compounds to be bioavailable in this way and transported across the blood brain barrier so readily – even glutathione needs to be taken intranasal for treatment of neurodegeneration.
Ergothioneine human trial: uptake metabolism and effects
While there is undeniably a lack of human trials exploring the in vivo effects of ERG, a recent study from 2017 explored ERG uptake, metabolism, and effects on biomarkers of oxidative damage and inflammation in healthy human subjects (cheah, Tang). One of the most interesting aspects of ERG discussed in this particular article is that ERG is a tautomer. This means that it exists in two forms – thione and thiol. The thiol is a single bond to sulfur and the thione is a double bond to sulfur. The article explains that in animal physiologic conditions, ERG primarily exists as the thione tautomer – under circumstances of low stress in the body, ERG remains in its thione tautomer form and is not the first choice as an antioxidant, rather endogenous antioxidants like glutathione are preferred (glutathione is a primary antioxidant thiol in the body). When the cells undergo higher levels of oxidative stress, ERG transforms into its thiol tautomer form and is then used for extra support. Additionally, under levels of elevated stress, tissues have increased amounts of the ERG, supposedly by upregulating expression of the OCTN1 ERG transporter in response to inflammatory cytokines.
This study also found that ERG can be stored in the cells for up to a month. It is theorized that ERG is stored for an extended period of time following consumption until it is required as a stronger defense mechanism.
During administration of ERG, plasma levels significantly elevated, while whole blood levels steadily increased for up to four weeks after administration stopped (red blood cells could continue to take ERG up as needed), and the excretion of ERG in the urine remained low, indicating that ERG is absorbed and retained in the body after oral administration. While this study used pure ERG, other studies previously mentioned (Heller) prove this same high bioavailability of ERG from dietary mushrooms, and so I will consider this as true for dietary ERG.
Mushroom analyses of ergothioneine: Fruiting body or mycelium? Gilled mushrooms or polypore mushrooms?
ERG was highest in the fruiting body of Pleurotus ostreatus and in the mycelium of Pleurotus eryngii. Generally, fruiting bodies and mycelium contained different amounts of ERG, with Pleurotus genus containing the highest amounts overall (Chen). Among simple mushrooms, fruiting body of White Button had the least (1.4mg erg per 85g mushroom) and Portabella the highest (2.7mg erg per 85g mushroom). While among specialty mushrooms, Maitake had the least (16.3mg erg per 85g mushroom) and oyster the highest (26.4mg per 85g mushroom) (Dubost).
Fruiting bodies of gilled mushrooms, specifically the cap, contain the most ERG, while polypore mushrooms contain very small amounts. Interestingly, the mycelium of Ganoderma contains more ERG than the fruiting body, but still not as much as the fruiting bodies of Shitake, Matsutake, Oyster, and Maitake (Lee, Wang).
Extraction:
Hot water extraction is sufficient to extract ERG from mushrooms.
In a study that explored water extraction – temperature of water, ratio of water to mycelium and the extraction rate of ERG, the following information was found:
Most notable: at 85°C and 20:1 ratio of mycelium to water, the extraction rate of ERG was 91.2% – the least effective yield of ERG was at 78°C and a 20:1 ratio of mycelium to water. There was no difference in ERG concentration between 5 minutes and 120 minutes, so a long boil to extract EGR is not necessary (Zhang).
In Summary:
Ergothioneine is a water soluble compound that is most abundant in Oyster mushrooms. There are transporters on different tissues in the body that are highly specific to ergothioneine. Ergothioneine is readily absorbed into the blood after consumption of mushrooms and stored in tissues for up to 1 month. In times of excessive oxidative stress, ergothioneine is taken up by those tissues and used as an antioxidant. Of note: there are transporters on the blood brain barrier and there is an association with low ergothioneine and age-related cognitive decline. In a world full of environmental toxins that are mostly impossible to escape, we might as well eat more mushrooms and get some extra protection.
How to apply this information to your life:
Eat a lot of mushrooms, especially Oyster mushrooms.
Antonicelli F, Aruoma OI. Ergothioneine inhibits oxidative stress- and TNF- a -induced NF- j B activation and interleukin-8 release in alveolar epithelial cells. 2003;302:860-864. doi:10.1016/S0006-291X(03)00224-9.
Aruoma OI, Spencer JPE, Mahmood N. Protection Against Oxidative Damage and Cell Death by the Natural Antioxidant Ergothioneine. 1999;37.
Aruoma OI, Whiteman M, England TG, Halliwell B. Antioxidant Action of Ergothioneine : Assessment of Its Ability to Scavenge Peroxynitrite. 1997;391(231):389-391.
Aubert DIRKT. Dietary Sources and Antioxidant Effects of Ergothioneine. 2007:6466-6474. doi:10.1021/jf071328f.
Benson KF, Ager DM, Landes B, Aruoma OI, Jensen GS. Improvement of joint range of motion ( ROM ) and reduction of chronic pain after consumption of an ergothioneine-containing nutritional supplement. Prev Med (Baltim). 2018;54(2012):S83-S89. doi:10.1016/j.ypmed.2012.02.001.
Betteridge, JD What Is Oxidative Stress? 2000:3-8.
Blankenfeldt W, Seebeck FP. Ergothioneine Biosynthetic Methyltransferase EgtD Reveals the Structural Basis of Aromatic Amino Acid Betaine Biosynthesis. 2015:119-125. doi:10.1002/cbic.201402522.
Cheah IK, Halliwell B. Biochimica et Biophysica Acta Ergothioneine ; antioxidant potential , physiological function and role in disease ☆. BBA – Mol Basis Dis. 2012;1822(5):784-793. doi:10.1016/j.bbadis.2011.09.017.
Chen S, Ho K, Hsieh Y, Wang L, Mau J. LWT – Food Science and Technology Contents of lovastatin , g -aminobutyric acid and ergothioneine in mushroom fruiting bodies and mycelia. LWT – Food Sci Technol. 2012;47(2):274-278. doi:10.1016/j.lwt.2012.01.019.
Deiana M, Rosa A, Casu V, et al. modulates oxidative damage in the kidney and liver of rats in vivo : studies upon the profile of polyunsaturated fatty acids L -Ergothioneine. 2004;5614:183-193. doi:10.1016/S0261-5614(03)00108-0.
Dubost NJ, Ou B, Beelman RB. Food Chemistry Quantification of polyphenols and ergothioneine in cultivated mushrooms and correlation to total antioxidant capacity. 2007;105:727-735. doi:10.1016/j.foodchem.2007.01.030.
Franzoni F, Colognato R, Galetta F, et al. An in vitro study on the free radical scavenging capacity of ergothioneine : comparison with reduced glutathione , uric acid and trolox. 2006;60:453-457. doi:10.1016/j.biopha.2006.07.015.
Gruber J, Fong S, Chen C, et al. Mitochondria-targeted antioxidants and metabolic modulators as pharmacological interventions to slow ageing. Biotechnol Adv. 2013;31(5):563-592. doi:10.1016/j.biotechadv.2012.09.005.
Gründemann D. The ergothioneine transporter controls and indicates ergothioneine activity — A review. Prev Med (Baltim). 2018;54(2012):S71-S74. doi:10.1016/j.ypmed.2011.12.001.
Halliwell B, Cheah IK, Drum CL. Biochemical and Biophysical Research Communications Ergothioneine , an adaptive antioxidant for the protection of injured tissues ? A hypothesis. Biochem Biophys Res Commun. 2016;470(2):245-250. doi:10.1016/j.bbrc.2015.12.124.
Harlfinger S, Golz S, Geerts A, et al. Discovery of the ergothioneine transporter. 2005.
Hseu Y, Lo H, Korivi M, Tsai Y, Tang M. Free Radical Biology and Medicine Dermato-protective properties of ergothioneine through induction of Nrf2 / ARE-mediated antioxidant genes in UVA-irradiated Human keratinocytes. Free Radic Biol Med. 2015;86(91):102-117. doi:10.1016/j.freeradbiomed.2015.05.026.
Kalaras MD, Richie JP, Calcagnotto A, Beelman RB. Mushrooms: A rich source of the antioxidants ergothioneine and glutathione. Food Chem. 2017;233:429-433. doi:10.1016/j.foodchem.2017.04.109.
Kerley RN, Mccarthy C, Kell DB, Kenny LC. Free Radical Biology and Medicine The potential therapeutic e ff ects of ergothioneine in pre-eclampsia. 2018;117(August 2017):145-157. doi:10.1016/j.freeradbiomed.2017.12.030.
Lee WY, Park E-J, Ahn JK, Ka K-H. Ergothioneine Contents in Fruiting Bodies and Their Enhancement in Mycelial Cultures by the Addition of Methionine. Mycobiology. 2009;37(1):43. doi:10.4489/MYCO.2009.37.1.043.
Lo Y, Lin S, Ulziijargal E, et al. Comparative Study of Contents of Several Bioactive Components in Fruiting Bodies and Mycelia of Culinary-Medicinal Mushrooms. 2012;14(4):357-363.
Li RWS, Yang C, Sit ASM, et al. Uptake and Protective Effects of Ergothioneine in Human Endothelial Cells. 2014;(September):691-700.
Markova NG, Karaman-jurukovska N, Dong KK, Damaghi N, Smiles KA, Yarosh DB. Free Radical Biology & Medicine Skin cells and tissue are capable of using L -ergothioneine as an integral component of their antioxidant defense system. Free Radic Biol Med. 2009;46(8):1168-1176. doi:10.1016/j.freeradbiomed.2009.01.021.
Martin KR. The Bioactive Agent Ergothioneine, a Key Component of Dietary Mushrooms, Inhibits Monocyte Binding to Endothelial Cells Characteristic of Early Cardiovascular Disease. 2010;13(6):1340-1346.
Moncaster JA, Walsh DT, Gentleman SM, Jen L, Aruoma OI. Ergothioneine treatment protects neurons against N -methyl- d – aspartate excitotoxicity in an in vivo rat retinal model. 2002;328:55-59.
Nakamichi N, Nakayama K, Ishimoto T, Masuo Y. Food-derived hydrophilic antioxidant ergothioneine is distributed to the brain and exerts antidepressant effect in mice. 2016;477:1-10. doi:10.1002/brb3.477.
Nguyen TH, Giri A, Ohshima T. A rapid HPLC post-column reaction analysis for the quantification of ergothioneine in edible mushrooms and in animals fed a diet supplemented with extracts from the processing waste of cultivated mushrooms. Food Chem. 2012;133(2):585-591. doi:10.1016/j.foodchem.2012.01.061.
Onofrio ND, Servillo L, Giovane A, et al. Free Radical Biology and Medicine Ergothioneine oxidation in the protection against high-glucose induced endothelial senescence : Involvement of SIRT1 and SIRT6. Free Radic Biol Med. 2016;96:211-222. doi:10.1016/j.freeradbiomed.2016.04.013.
Paul, BD, Snyder, SH. The Unusual Amino Acid, l-ergothioneine is a Physiologic Cytoprotectant. NIH public access 2010: 17-7. Doi: 10.1038/cdd.2009.163
Sakrak O, Kerem M, Bedirli A, et al. Ergothioneine Modulates Proinflammatory Cytokines and Heat Shock Protein 70 in Mesenteric Ischemia and Reperfusion Injury. 2018;42(2008):36-42. doi:10.1016/j.jss.2007.04.020.
Servillo L, Castaldo D, Casale R, et al. Free Radical Biology and Medicine An uncommon redox behavior sheds light on the cellular antioxidant properties of ergothioneine. Free Radic Biol Med. 2015;79:228-236. doi:10.1016/j.freeradbiomed.2014.11.017.
Shah SP, Duda JE. Dietary modifications in Parkinson ’ s disease : A neuroprotective intervention ? Med Hypotheses. 2018;85(6):1002-1005. doi:10.1016/j.mehy.2015.08.018.
Song T, Chen C, Liao J, Ou H, Tsai M. Ergothioneine protects against neuronal injury induced by cisplatin both in vitro and in vivo. Food Chem Toxicol. 2010;48(12):3492-3499. doi:10.1016/j.fct.2010.09.030.
Tomomi I, Manami K, Hironobu T, Etsuko H, Toshio N, Toshihiko O. Ergothioneine as an Anti-Oxidative / Anti-Inflammatory Component in Several Edible Mushrooms. 2011;17(2):103-110.
Wang X, Zhang J, Wu L, et al. A mini-review of chemical composition and nutritional value of edible wild-grown mushroom from China. Food Chem. 2014;151:279-285. doi:10.1016/j.foodchem.2013.11.062.
Weigand-heller AJ, Kris-etherton PM, Beelman RB. The bioavailability of ergothioneine from mushrooms ( Agaricus bisporus ) and the acute effects on antioxidant capacity and biomarkers of in fl ammation. Prev Med (Baltim). 2018;54(2012):S75-S78. doi:10.1016/j.ypmed.2011.12.028.
Wei Z, Li X, Li X, Liu Q, Cheng Y. Oxidative Stress in Parkinson ’ s Disease : A Systematic Review and. 2018;11(July):1-7. doi:10.3389/fnmol.2018.00236.
Yoshida S, Shime H, Funami K, Takaki H. The Anti-Oxidant Ergothioneine Augments the Immunomodulatory Function of TLR Agonists by Direct Action on Macrophages. 2017:1-15. doi:10.1371/journal.pone.0169360.